◄ Carnets Geol. 19 (18) ►
![]()
Outline:
[1. Introduction]
[2. Geographical and geological settings and material]
[3. General features of the intergrowth]
[4. Cases of immuration]
[5. Systematic palaeontology]
[6. Evaluation of the association]
[7. Conclusions]
and ...
[Bibliographic references]
Université de Bretagne Occidentale, Laboratoire de Paléontologie, UFR Sciences & Techniques, 6 avenue Le Gorgeu, CS 93837, F-29283, Brest (France)
Muséum National d'Histoire Naturelle, Département Origines et Évolution CP 38, 8 rue de Buffon, 75005, Paris (France)
Published online in final form (pdf) on November 11, 2019
DOI 10.4267/2042/70538
![]()
[Editor: Bruno Granier;
language editor: Stephen Eager]
![]()
Numerous and generally well-preserved examples of the intergrowth Rugosa-Bryozoa from the Upper Pragian and Lower Emsian of the Armorican Massif (Châteaulin and Laval synclinoria), France, and from the Upper Emsian of the Ougarta Mountains, Erg Djemel, Algeria, are described. In the Armorican Massif, the corallites of a rugosan Tryplasmatidae? are intergrown with Ceramoporidae bryozoan close to Crepipora, exceptionally with an unidentified Fistuliporidae (likely a new genus), whereas in Ougarta the coral is not identifiable and is associated with a Fistuliporidae assigned to Fistulipora. Although mainly left in open nomenclature, the material is fully described (structure and microstructure) and illustrated (calcitic skeleton and natural moulds) for the first time. In addition, the presence of Ceramoporidae in the Lower Devonian is clearly established. The evaluation of the association is briefly discussed and a mutualistic relationship supported.
intergrowth;
Rugosa;
Bryozoa;
Pragian-Emsian;
Armorican Massif;
Algerian Sahara
Plusquellec Y. & Bigey F.P. (2019).- New data on the intergrowth of Rugosa-Bryozoa in the Lower Devonian of North Gondwana.- Carnets Geol., Madrid, vol. 19, no. 18, p. 421-437.
Données nouvelles sur l'intercroissance Rugosa - Bryozoa dans le Dévonien inférieur du Nord Gondwana.- Des exemples nombreux et généralement bien conservés d'intercroissance Bryozoa-Rugosa, collectés dans le Praguien supérieur et l'Emsien inférieur du Massif Armoricain (synclinoria de Châteaulin et de Laval), France, et dans l'Emsien supérieur des Monts d'Ougarta, Erg Djemel, Algérie, sont décrits. Dans le Massif Armoricain, les corallites d'un tétracoralliaire Tryplasmatidae? sont associés à un bryozoaire Ceramoporidae proche de Crepipora, exceptionnellement à un Fistuliporidae probablement nouveau, tandis que dans l'Ougarta le coralliaire - mal ou partiellement préservé - n'est pas identifiable et se développe en association avec un Fistuliporidae du genre Fistulipora. Bien que laissé en nomenclature ouverte pour l'essentiel, le matériel est décrit en détail (structure et microstructure) et, des images du squelette calcitique et des moulages naturels sont fournies pour la première fois. Accessoirement, la présence des Ceramoporidae dans le dévonien inférieur est incontestablement établie. Les modalités de l'intercroissance sont brièvement discutées et une relation de type mutualiste est retenue.
intercroissance ;
Rugosa ;
Bryozoa ;
Praguien-Emsien ;
Massif Armoricain ;
Sahara Algérien
Vinn et al. (2016, 2017a, 2018) describe some cases of intergrowth between Rugosa and Bryozoa in the Katian of Estonia and stated that this association is only known from the Upper Ordovician of Baltica and Laurussia. Shortly after, Sendino et al. (2019) publish a "new rugose coral-cystoporata association from the Devonian of NW Spain" and stated that it is "the first evidence of intergrowths between Devonian rugose corals and bryozoans". In fact some cases of intergrowths were previously reported in an abstract published on the occasion of the Congrès national de Paléontologie held in Paris in 1990 (Plusquellec & Bigey, 1990), but not illustrated and the aim of this work is to provide new accurate data about these cases known from the Devonian of the Armoricain Massif (France) and the Ougarta Mountains (Algeria).
As far as we know, no other example of rugose coral - bryozoan intergrowths have been published. However, a similar association involving the bryozoan Celleporaria palmata (Michelin, 1847) and the scleractinian corals referred either to Culicia parasitica (Michelin, 1847) or Cryptangia woodii Milne-Edwards & Haime, 1850, is well known from the Neogene of northwestern Europe (Cadée & McKinney, 1994).
The Devonian material described in this paper has been collected in four localities of the Armorican Massif belonging to the Central Armorican domain and in one locality of the Ougarta area. These two areas are situated during the Devonian time on the northern margin of the paleocontinent Gondwana.
In the Western part of the Armorican Massif, Châteaulin Synclinorium, three fossiliferous localities, located in the rade de Brest and Crozon peninsula, have yielded the association.
1 - Pointe de l'Armorique, Northern section, West of Plougastel-Daoulas (Finistère), upper part of the Le Faou Formation in the level called "Banc des monstres", Lower Emsian, dehiscens Conodont Zone (six specimens in natural moulds, LPB 17 257 - 17 258.a-e).
2 - Seillou section, left bank of the Le Faou River (Finistère), upper part of the Le Faou Formation, level LF 7 sensu Le Menn (1985), "Banc des monstres", Lower Emsian, dehiscens Conodont Zone (one specimen in natural mould, LPB 17 259).
3 - Batterie de Pont-Scorff section, Eastern coast of Roscanvel area, Crozon peninsula (Finistère), calcareous level in the Le Faou Formation, Upper Pragian or Lower Emsian (one specimen with preserved skeleton, LPB 17 256).
In the Eastern part of the Armorican Massif, Laval Synclinorium, old quarries located in the NW of Loué (Sarthe) known as "carrières de la station de pompage de Monfoulon" (Plusquellec et al., 1993, fig. 22) have yielded very well-preserved specimens with calcitic skeletal material in an horizon called Monfoulon Limestones, roughly middle part of the Montguyon Formation. The material was collected from two levels, respectively 4.50 m ("niveau à Tabulés branchus") and 10 m ("niveau à Blastoïdes") above a level which has provided the Upper Pragian conodont Polygnathus pireneae. (Plusquellec et al., 1993, p. 28, fig. 22). Caudicriodus angustoides angustoides and Caudicriodus curvicauda are known from two levels, one between the "niveau à Tabulés branchus" and the "niveau à Blastoïdes", and a second one about 10 m above the "niveau à Blastoïdes" (Weyant and Morzadec unpublished data). According to Weyant et al. (2010), C. curvicauda occurs well below the Lower Emsian P. dehiscens in the Guadarrama and in the Upper part of the Pragian in different sections from the Barrandian area.
Thus, in the quarry of Monfoulon, the Rugosa-Bryozoa intergrowths are Upper Pragian (pireneae Zone) in age, including the following material: one specimen probably from the "niveau à Tabulés branchus", LPB 17 252; five specimens from the "niveau à Blastoïdes", LPB 17 248.a-b, 17 249, 17 250, 17 253, 17 254 and two specimens for which the level is unknown, LPB 17 251 and 17 259bis.
In the Ougarta Mountains, the material is from the Erg Djemel section, Chefar El Ahmar Formation, level ED 29 ("niveau coralligène"), Upper Emsian (Paris et al., 1997). One specimen with preserved skeleton, LPB 17 255.
As a result, the material consists in 18 specimens, 35 thin sections (including 13 polished ultra-thin sections, the so-called "LFP"), 46 acetate peels (including 30 serial acetate peels in LPB 17 249) and is deposited in the collections of the Laboratoire de Paléontologie, Université de Bretagne Occidentale, Brest (France), with the prefix LPB.
The rugosan corallites, tentatively assigned to the Tryplasmatidae, occur embedded in bryozoan belonging to n.gen. 1 cf. Crepipora Ulrich, 1882 (Cystoporata, Ceramoporidae), Fistulipora McCoy, 1849 (Cystoporata, Fistuliporidae), and to a third unidentified genus (n.gen. 2) of the family Fistuliporidae.
The association with the Crepipora-like is the most frequent
(Figs. 1.A ![]() ,
2
,
2 ![]() -
3
-
3 ![]() ) and is recorded
in the material from Monfoulon: LPB 17 248 - 252, LPB 17 253 ?, 17 259bis
and from the Batterie de Pont-Scorff: LPB 17 256. It is very likely that
the specimens preserved in natural moulds (Pointe de l'Armorique and Seillou)
belongs to the same association owing to the characteristics of the apparently
contiguous moulds of autozooecia: LPB 17 257 - 259. The association
with the Fistuliporidae n.gen. 2 is only known in Monfoulon: LPB 17 254 (Figs.
12.C-D
) and is recorded
in the material from Monfoulon: LPB 17 248 - 252, LPB 17 253 ?, 17 259bis
and from the Batterie de Pont-Scorff: LPB 17 256. It is very likely that
the specimens preserved in natural moulds (Pointe de l'Armorique and Seillou)
belongs to the same association owing to the characteristics of the apparently
contiguous moulds of autozooecia: LPB 17 257 - 259. The association
with the Fistuliporidae n.gen. 2 is only known in Monfoulon: LPB 17 254 (Figs.
12.C-D ![]() ,
14
,
14 ![]() ), the one involving Fistulipora
in Ougarta: LPB 17 255 (Figs. 4
), the one involving Fistulipora
in Ougarta: LPB 17 255 (Figs. 4 ![]() - 5
- 5 ![]() ).
).
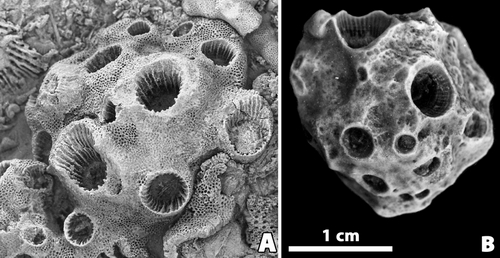
The zooarium of all the bryozoan material is massive
and more or less hemispherical. In the specimen from Ougarta the proximal side
of the colony is rather well preserved: concave with concentric growth ridges
and no indication of the presence of coral. The distal side is generally
irregularly convex and shows the numerous zooecial apertures as well as the
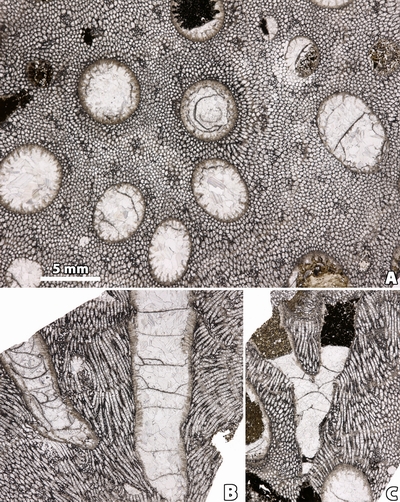
calical apertures of the associated coral (Fig. 1 ![]() ).
).
The rugosan corallites are regularly spaced over the colony surface leaving only their calical aperture free. Moreover, although coated by the zooecia on their external side, the apertures of the corallites are elevated above the growth surface of the bryozoan. The size of the corallites at the surface level is highly variable indicating various growth stages. In addition, the longitudinal sections as well as the natural moulds show that the corals began to grow after the bryozoan became established on the substrate.
As in the rugosan-stromatoporoid association (Vinn et al., 2015) and rugosan-tabulate association (Vinn et al., 2017b) the rugosan corallites are arranged perpendicularly to the growth surface of the bryozoan. The longitudinal section in the association shows that the bryozoan grew by frontal budding and according to Cadée and McKinney (1994) "the growth of the corallites was nicely balanced, keeping the corallites always at the colony surface".
In
longitudinal section, around each corallite, the zooecia appear upwards
diverging with an angle of about 25-45° and the virtual laminations within
the bryozoan (shown by the diaphragms) are upturned when they meet corallites (Figs.
2.B ![]() ,
3.A, 3.C
,
3.A, 3.C ![]() ).
).
Frequently, it can be seen that, at the contact between the
bryozoan and the coral, the wall of the zooecia is lacking (Figs.
3 ![]() ,
12.A
,
12.A ![]() ). This feature is seen in
both transverse and longitudinal sections and
indicates that the growth of the coral precedes that of the bryozoan. Similar
features have been described and well-illustrated by Sorauf and Kissling
(2012) in a Streptelasma anchored
within the skeleton of Paleofavosites:
usually, the ventral wall of this solitary Rugosa is absent or very thin.
). This feature is seen in
both transverse and longitudinal sections and
indicates that the growth of the coral precedes that of the bryozoan. Similar
features have been described and well-illustrated by Sorauf and Kissling
(2012) in a Streptelasma anchored
within the skeleton of Paleofavosites:
usually, the ventral wall of this solitary Rugosa is absent or very thin.
Rarely, necroses of the surface are recorded within the
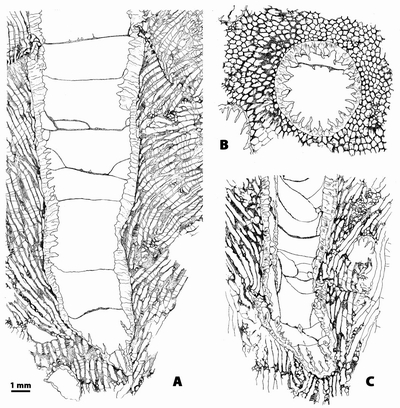
bryozoans; in the illustrated example it is located around the calicinal edge
of the Rugosa (Fig. 3.A ![]() ).
).
Despite the appearances the
intergrowth occurs between the bryozoan and solitary corals. The branching
pattern is widely missing as shown by transverse and longitudinal sections,
and also by the natural moulds. A unique pattern of parricidal increase has been
noted, where an adult corallite split into two offsets of the same diameter
occupying the whole calice of the parent (LPB 17 250, thin section b, Fig.
2.C ![]() ). It is obvious too, taking into account the various sizes and length of the
corallites and the very scarce case of budding, that the settlement of the
young corals on the surface of the bryozoan was an ongoing process.
). It is obvious too, taking into account the various sizes and length of the
corallites and the very scarce case of budding, that the settlement of the
young corals on the surface of the bryozoan was an ongoing process.
As a result, the association involves a bryozoan and a gregarious solitary coral giving rise - due to the "cement" provided by the bryozoan - to a pseudo-colony of Rugosa. The collected material does not allow us to say if the bryozoan occurs with or without the coral (but it is very likely) neither if the coral can be found solitary.
Click on thumbnail to enlarge the image. |
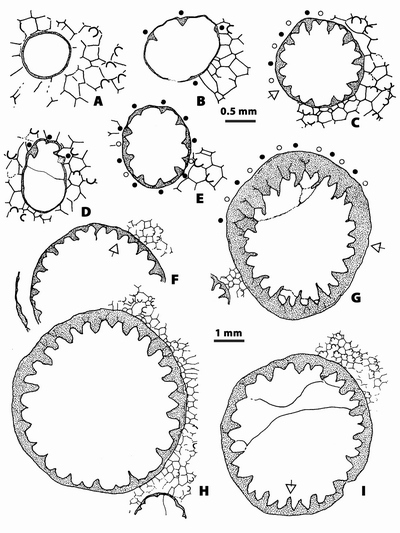
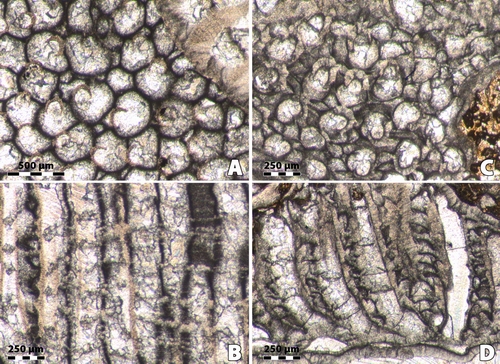
Figure 1:
A,
Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth, distal side; LPB 17 248.
Monfoulon, Armorican Massif, Monfoulon Limestones, Upper Pragian. B,
Fistulipora sp. - Rugosa indet.
intergrowth, distal side; LPB 17 255. Ougarta, Algeria, Erg Djemel section,
Chefar El Ahmar Fm., Upper Emsian. |
|
Figure
2:
A, Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth. A,
transverse section, note in the Bryozoa the numerous maculae; LPB 17 251,
thin section a. B, longitudinal
section; LPB 17 251, thin section b. C,
longitudinal section showing a case of parricidal increase; LPB 17 250,
thin section b. Monfoulon, Armorican Massif, Monfoulon Limestones, Upper Pragian. |
|
Figure
3: A,
Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth. A,
longitudinal section, note in the upper part of the section a surface of
necrosis in the bryozoan linked with irregularities in the growing of the coral;
LPB 17 251, thin section b. B,
transverse section; LPB 17 251, thin section a. C, longitudinal section, LPB 17 251, thin section b.
Monfoulon,
Armorican Massif, Monfoulon Limestones, Upper Pragian-Lower Emsian. |
|
Figure
4:
Fistulipora
sp. - Rugosa indet. intergrowth. A,
transverse section in the Rugosa, oblique in the Bryozoa. B, transverse section; both LPB 17 255, thin section B 35 142-a
(same specimen as Figs. 1.B |
|
Figure
5:
Fistulipora
sp. - Rugosa indet. intergrowth (same specimen as Fig. 1.B |
In addition to the intergrowth, some corallites are
found to be overgrown by the bryozoan in specimens from Monfoulon (LPB 17 250,
LPB 17 254, LPB 17 259bis) and Pont-Scorff (LPB 17 256, Fig. 6 ![]() ).
).
This may be caused by active overgrowth but more likely, passively after death of the corallite (cf. Kershaw, 1987). According to Sorauf and Kissling (2012), dealing with the immuration of Streptelasma by Paleofavosites, the death of the corallite appears to have preceded overgrowth as the calices of some specimens seem to be filled with sediments prior to be immured. Moreover, overgrowth only occurs on corallites reaching a large diameter, i.e., adult or gerontic specimens.
The overgrowth is either located at the calicinal
margin level or spread more or less deeply on its inner side. In the specimen
from Pont-Scorff, the inner side of the calice is occupied by a "foreign"
bryozoan (Fistulipora sp.) prior to
be overlapped by its "own" Crepipora-like (Fig. 6 ![]() ).
).
|
Figure
6:
Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth overgrown by Fistulipora sp., longitudinal section; loose hatching as Crepipora,
close hatching as Fistulipora, X as
(late) boring; LPB 17 256, thin section B 35 145. Pont-Scorff,
Roscanvel peninsula, Armorican Massif, Le Faou Fm., Upper Pragian-Lower Emsian. |
Subclass Rugosa Milne-Edwards & Haime, 1850
Order Cystiphyllidae Nicholson, 1889
Family ?Tryplasmatidae Etheridge, 1907
Genus ?n.gen.
See discussion below.
n.gen.? n.sp. 1
Morphology. Corallum solitary or with very scarce
parricidal offsets (Fig. 2.C ![]() ); corallites ceratoid to cylindrical. Apical
region conical, strongly curved to geniculate in some cases (Fig. 7
); corallites ceratoid to cylindrical. Apical
region conical, strongly curved to geniculate in some cases (Fig. 7 ![]() )
and
devoid of septa (Fig. 8.A
)
and
devoid of septa (Fig. 8.A ![]() ) but "ornamented" by delicate transverse growth
striae visible on the natural cast of the inside of corallite. This unusual feature
shows some similarity with that
illustrated (on the external surface) by Fedorowski
(1991) in Paraduplophyllum. Calices having carinate septa differentiated in
two orders (Fig. 8.C-I
) but "ornamented" by delicate transverse growth
striae visible on the natural cast of the inside of corallite. This unusual feature
shows some similarity with that
illustrated (on the external surface) by Fedorowski
(1991) in Paraduplophyllum. Calices having carinate septa differentiated in
two orders (Fig. 8.C-I ![]() ). External surface not exposed due to the intergrowth.
). External surface not exposed due to the intergrowth.
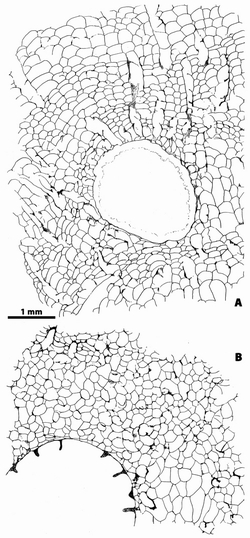
Transverse sections. In the juveniles sections, close
to the aseptate apical region, only few septa are recorded (3 to 4). They are
short and likely major ones (Fig. 8.B, 8.D ![]() ), whereas in the adult or gerontic
sections the number of septa reaches up
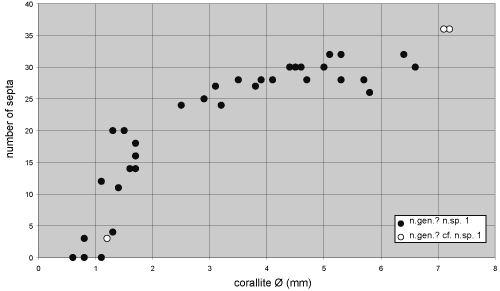
to 32 (major and minor septa). In fact, the number of septa increases quickly but does not
exceed 20 in the corallites the diameter of which is less than 2 mm, whereas
the number only varies from 24 to 32 in the corallites the
diameter of which is between 2 and 7 mm (Diagram 1
), whereas in the adult or gerontic
sections the number of septa reaches up
to 32 (major and minor septa). In fact, the number of septa increases quickly but does not
exceed 20 in the corallites the diameter of which is less than 2 mm, whereas
the number only varies from 24 to 32 in the corallites the
diameter of which is between 2 and 7 mm (Diagram 1 ![]() ).
).
|
The protosepta cannot be identified although the
sections (especially the adult sections) generally show an area where major
and minor septa are less prominent, less thick, and in some cases difficult to
distinguish from each other (Fig. 8.C, 8.F-G, 8.I ![]() ).
).
Overall, the major septa are short, thicker than the
minor ones, with an inner margin rather smooth or bearing a small tooth (Fig.
8.I ![]() ). In some sections, the outline of the lateral side of the septa appears
irregular due to the presence of carinae (Fig. 8.G
). In some sections, the outline of the lateral side of the septa appears
irregular due to the presence of carinae (Fig. 8.G ![]() ). The minor septa are
shorter and usually triangular in section.
). The minor septa are
shorter and usually triangular in section.
Longitudinal sections. The tabulae, thin, mainly flat,
appear widely spaced, generally between 0.4 to 2.8 mm, some being as much as
3.6 to 4.2 mm, even 6.8 mm apart. Rare tabulae show slight thickening and bear
short spines. Some incomplete, convex and rather deeply inclined peripheral
tabellae are recorded; dissepiments are absents (Fig. 3.A, 3.C ![]() ).
).
Microstructure. The septa exhibit a trabecular
microstructure consisting of parallel and contiguous rather short bundles of
fibres, free at inner ends, sloping upwards and inwards from the periphery at
angle ca. 25° (Figs. 3.A, 3.C ![]() , 9
, 9 ![]() ).
The trabeculae are devoid of true axis as in the monacanths and thus are
assigned to water-jet trabeculae (see Cuif & Gautret,
1993,
fig. 4).
).
The trabeculae are devoid of true axis as in the monacanths and thus are
assigned to water-jet trabeculae (see Cuif & Gautret,
1993,
fig. 4).
The wall (studied as well as the septa by ultra-thin sections, the so-called "LFP") consists of two parts:
1) a very thin outer layer (0.05 - 0.07 mm) of fibroids
not organized into trabeculae and oriented inwards and upwards (=true epitheca?)
(Fig. 9.B-C ![]() ). A similar structure is known in Siphonodendron and
probably in Disphyllum
(see respectively Semenoff-Tian-Chansky, 1984, text-fig. 2F, and Coen-Aubert
et al., 2013, fig. 7).
). A similar structure is known in Siphonodendron and
probably in Disphyllum
(see respectively Semenoff-Tian-Chansky, 1984, text-fig. 2F, and Coen-Aubert
et al., 2013, fig. 7).
2) an inner thick layer (0.2 - 0.3 mm) built of the
thickened outer parts of the radial elements (major and minor septa) (Fig.
9.A ![]() ). In addition, supernumerary bundles of fibres occupy spaces between major
and minor septa but are confined to the stereozone although their inner margin
- seen in transverse section - could be slightly prominent (Fig.
9.A
). In addition, supernumerary bundles of fibres occupy spaces between major
and minor septa but are confined to the stereozone although their inner margin
- seen in transverse section - could be slightly prominent (Fig.
9.A ![]() ).
Thus, the wall seems to be better interpreted as trabeculotheca (or a
structure close to this category) than to septotheca (see Stolarsky,
1996, fig. 7).
).
Thus, the wall seems to be better interpreted as trabeculotheca (or a
structure close to this category) than to septotheca (see Stolarsky,
1996, fig. 7).
|
Figure
7:
Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth preserved in natural moulds,
note the curved tip of the corallites and the development of transverse striae;
LPB 17 258.e. Pointe
de l'Armorique section, W. of Plougastel-Daoulas, Armorican Massif, Le Faou
Fm., "Banc des monstres", Lower Emsian. |
|
Figure
8:
Bryozoa (n.gen. 1 cf. Crepipora
n.sp. 1).- Rugosa (n.gen.? n.sp. 1) intergrowth. Transverse sections in the
corallites at various growth stages. Black circle as major septum, open circle
as minor septum, open arrow as area where major and minor are few
differentiated. Note the lack of septa in earliest stage (A)
and only three or four major septa in juvenile stages (B, D). A,
LPB 17 259 bis, thin section B 36 854. B, LPB 17 259 bis, thin section B 36 854. C,
LPB 17 254, thin section a. D,
LPB 17 252, thin section a. E,
LPB 17 252, thin section c. F,
LPB 17 252, thin section c. G,
LPB 17 250, thin section b. H,
LPB 17 259 bis, thin section B 36 854. I, note a small tooth on the margin of the major septum situated on
top of figure, LPB 17 252, thin section a. Monfoulon, Armorican Massif,
Monfoulon Limestones, Upper Pragian. |
|
Figure
9:
Rugosa Tryplasmatidae?, n.gen.? n.sp. 1. Microstructure of the wall and septa. Black
circle as major septum, open circle as minor septum, open square as
supernumerary bundles of fibres, "olf" as outer layer of fibroids. A,
transverse section; LPB 17 250, thin section a (standard thickness). B,
idem; LPB 17 250, "LFP" thin section l. C, longitudinal section within the plane of septa; LPB 17 250,
"LFP" thin section i. D, longitudinal section at right angle to the plane of septa showing how the
trabeculae give rise to carinae; LPB 17 259 bis, thin section B 36 856
(standard thickness). Monfoulon, Armorican Massif, Monfoulon Limestones, Upper
Pragian. |
Remark. One specimen from Monfoulon (LPB 17 253),
although associated with Crepipora,
differs from the material described above, only by having longer major septa
in all the stages of growth (Fig. 10 ![]() ,
Diagram 1
,
Diagram 1 ![]() ); it is provisionally assigned to
n.gen.? cf. n.sp. 1. As for that of Ougarta (Figs. 1.B
); it is provisionally assigned to
n.gen.? cf. n.sp. 1. As for that of Ougarta (Figs. 1.B ![]() , 4
, 4 ![]() - 5
- 5 ![]() ), very poorly
preserved in some sections, even wall possibly lacking (Fig. 5.A
), very poorly
preserved in some sections, even wall possibly lacking (Fig. 5.A ![]() ), the coral remains
unidentified; it is associated with Fistulipora.
), the coral remains
unidentified; it is associated with Fistulipora.
Discussion. The material shares some generic features with Tryplasma Lonsdale, 1845 (see diagnosis in Hill, 1981, p. F98), but according to Pedder (pers. comm., 1990) it probably represents a new genus and the family assignment is also problematical.
It is obvious that, by the use of "LFP", the level of knowledge of our specimens is better than that of Tryplasma (description of layer of fibroids, supernumerary bundles of fibres in the wall) and thus the comparison is uncertain as such details are not available for Tryplasma. Moreover, the septa of Tryplasma are said to be rhabdacanthine, holacanthine or dimorphacanthine - strange in our opinion - whereas the Armorican material is undoubtedly water-jet.
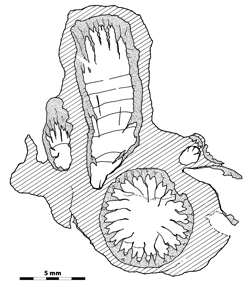
|
Figure
10:
Bryozoa (n.gen. 1 cf. Crepipora n.sp. 1).-
Rugosa (n.gen.? cf. n.sp. 1) intergrowth, note that the septa are longer than
usual in the other specimens of this locality; note also a juvenile section with
three major septa located in one side of the section. LPB 17 253, thin
section a. Monfoulon, Armorican Massif, Monfoulon Limestones, Upper Pragian. |
Preliminary remarks about the generic assignment of the Bryozoa
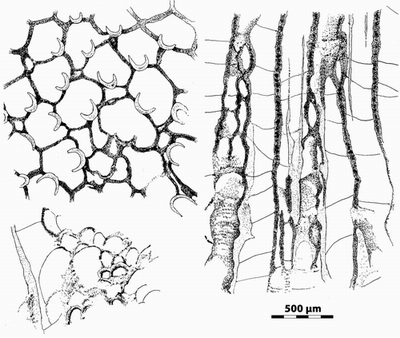
Case 1 - In the Treatise on Invertebrate Paleontology, Bryozoa revised, Boardman et al. (1983) stated that Ceramoporina primarily have a laminated skeletal microstructure (p. 342) and show the presence of "tabular crystallites" in the Ordovician Ceramophylla vaupeli (fig. 149) and Ceramoporella flabellata (fig. 150). They also provide numerous and nice photos, at lower magnification, showing the lamellar nature of most of the genera of the family such as Acanthoporella (fig. 157), Ceramophylla (fig. 158), Ceramoporella (fig. 159) and Crepipora (fig. 160), all from Ordovician.
These authors give less clear data about the microstructure of the lunaria deposits but pointed out their light colored hyaline appearance under a light microscope and indistinct, distantly spaced laminations in some forms (p. 344).
On the other hand, they did not indicate the occurrence of Ceramoporidae in the Devonian except that of the poorly known genus Ganiella whose "microstructure (is) indistinct, questionably laminated" and "the lunaria with shorter radius of curvature, indistinct" (p. 368). They did not mention the very poorly illustrated species of Crepipora ferganensis Orlovsky, 1982, and C. subglobosa Orlovsky, 1982, from the Lower Devonian of Asia (Modzalevskaja & Orlovsky, 1968).
Ultra-thin sections ("LFP") prepared from the
specimens LPB 17 250 and 17 251 from Monfoulon and formerly assigned
to Crepipora (Plusquellec
& Bigey, 1990) show that the wall of the autozoecia is made of
short and rather strong fibres 12-13 µm x 3 µm) displayed on both sides of a
median plate (Fig. 11.B, 11.C left ![]() ). This plate is made of tiny granular
crystals (3 µm), appearing as a dark line in transverse sections of usual
thickness (Fig. 11.A
). This plate is made of tiny granular
crystals (3 µm), appearing as a dark line in transverse sections of usual
thickness (Fig. 11.A ![]() ). In transverse section the fibres appear normal to the
median line, whereas in longitudinal one they show their slightly upwards
diverging setting (Fig. 11.C left
). In transverse section the fibres appear normal to the
median line, whereas in longitudinal one they show their slightly upwards
diverging setting (Fig. 11.C left ![]() ). The microstructure of the
lunaria, well
exposed in longitudinal sections, consists of large fibroids radiating upward
and outward (Fig. 11.C right
). The microstructure of the
lunaria, well
exposed in longitudinal sections, consists of large fibroids radiating upward
and outward (Fig. 11.C right ![]() ). The size of these crystallites (40-100 µm x
5-10 µm) gives rise to their light coloured appearance in sections of usual
thickness. In transverse section, the main visible feature of the lunaria (in
natural light) is the presence of more or less scattered tiny dark granules
such as those described in the large biocrystals of the tabulate coral Thamnopora by Lafuste and Tourneur
(1991).
). The size of these crystallites (40-100 µm x
5-10 µm) gives rise to their light coloured appearance in sections of usual
thickness. In transverse section, the main visible feature of the lunaria (in
natural light) is the presence of more or less scattered tiny dark granules
such as those described in the large biocrystals of the tabulate coral Thamnopora by Lafuste and Tourneur
(1991).
Hence, it appears that the microstructure of the autozooecial wall of the Devonian Crepipora-like forms clearly differs from that of the Ordovician forms. In addition, taking into account the light-coloured appearance of the lunaria of the Ordovician genera their microstructure could be similar to that of the Devonian representatives.
As a result, their microstructural characteristics indicate that the Devonian Crepipora-like forms belong to a new genus and they are herein provisionally assigned to n.gen. 1 cf. Crepipora Ulrich, 1882.
Remarks: Spjeldneas (1963, p. 66), dealing with the type species of Crepipora, indicated that "the pores are numerous in the exozone"; these are not recorded in the Devonian material.
Moreover, it can be noted that the evolution of the skeleton of the Ceramoporidae from Middle Ordovician to Lower Devonian is somewhat similar to that of the tabulate corals. For example, as early as 1962, Lafuste pointed out that the wall of the genus Favosites is microlamellar in Silurian forms but fibrous in Devonian ones, and that, accordingly, it should be split into two distinct genera.
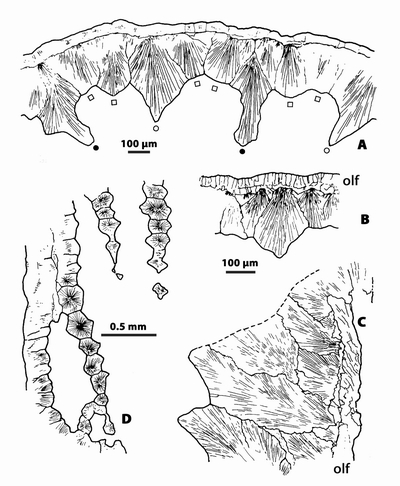
|
Figure
11:
n.gen. 1
cf. Crepipora n.sp. 1. Microstructure
of specimen LPB 17 250. A,
transverse section showing the median dark line of the autozoecial wall; note
that this feature is only visible rarely in some areas of the section; thin
section LPB 17 250-b (standard thickness). B, transverse section in the autozoecial wall, explanation in text;
"LFP" thin section 17 250-l. C,
longitudinal section in the autozoecial wall (left) and in the lunaria (right),
explanation in text; "LFP" thin section 17 250-h. D, longitudinal section in a lunaria showing the morphology of the
fibroids; "LFP" thin section 17 250-k. Monfoulon, Armorican Massif,
Monfoulon Limestones, Upper Pragian. |
Case 2 - Another member of the Cystoporata is recorded
from the Monfoulon Limestones in association with a species of the Rugosa. The
only available specimen is small and only a section of petrographic thickness
was possible. It is characterized by: 1) strongly thickened lunaria probably
made of fibroids (light coloured appearance) and 2) by well-developed
extrazooidal vesicular tissue. The microstructure of the wall of the
autozooecia is indistinct (owing to the lack of "LFP") whereas the alleged
fibroids of the lunaria are strongly oblique to the growth direction of the
lunaria itself, parallel to each other, and upwards sloping from a dark line
shifted close to their inner (concave) side (Fig. 14 ![]() ).
).
The specimen clearly belongs to the Fistuliporidae and differs from all genera of the family by its significantly thickened and strongly curved lunaria. Thus it is herein provisionally assigned to n.gen. 2. It recalls Buskopora Ulrich, 1886.
In addition, note that the microstructure of the lunaria of n.gen. 2 differs from that of n.gen. 1 cf. Crepipora and, as a result, examples of two kind of lunarial microstructure are provided: "water jet-like" (in sections normal to the flattening of the lunaria) in some Ceramoporidae, clinogonally fibrous in some Fistuliporidea.
Case 3 - The specimen from Ougarta is assigned to the large genus Fistulipora McCoy, 1849.
The recorded Bryozoa are thus as listed below.
Class Gymnolaemata Allman, 1856
Order Cystoporata Astrova, 1964
Family Ceramoporidae Ulrich, 1882
Genus n.gen. 1 cf. Crepipora Ulrich, 1882
n.gen. 1 cf. Crepipora n.sp. 1
(Figs. 1.A ![]() ,
2
,
2 ![]() -
3
-
3 ![]() ,
12.A-B
,
12.A-B ![]() ,
13
,
13 ![]() )
)
Remark: The species is not described herein but, owing to the lack of dependable data about the so-called Devonian Crepipora, it is likely a new species. Its description is in progress.
Family Fistuliporidae Ulrich, 1882
Genus Fistulipora McCoy, 1849
Fistulipora sp.
Remark: The only known specimen is not described but illustrated.
Genus n.gen. 2
n.gen. 2 n.sp. 2
(Fig. 12.C-D ![]() , 14
, 14 ![]() )
)
Remark: The well preserved but unique specimen is provisionally not described but illustrated.
|
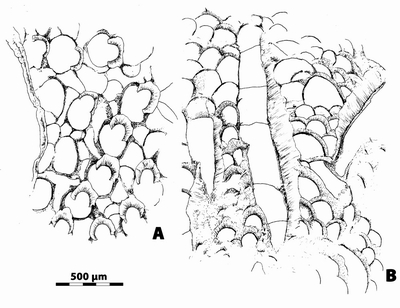
Figure
12:
A-B,
n.gen. 1 cf. Crepipora n.sp. 1,
respectively transverse and longitudinal section (same specimen as Fig. 13 |
|
Figure
13:
n.gen. 1
cf. Crepipora n.sp. 1. A,
transverse section showing the well- developed lunaria clearly projecting into
autozooecial cavity; LPB 17 251, thin section a. B,
longitudinal section showing long lunarial deposit (in light grey) reaching
herein up to 1.2 mm, and numerous diaphragms; LPB 17 251, thin section b. C, longitudinal section, lunarial deposit on left and thickened
vesicular heterozooecia on bottom and right; LPB 17 251, thin section c.
Monfoulon, Armorican Massif, Monfoulon Limestones, Upper Pragian. |
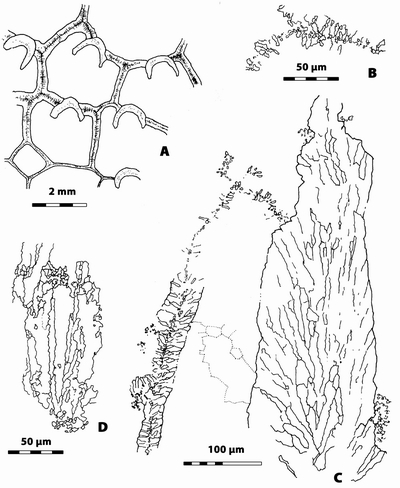
|
Figure
14:
n.gen. 2 n.sp. 2. A, transverse section showing the strongly thickened lunaria and, on
left, section in the wall of an indet. Rugosa (note the presence of a median
dark line not recorded in Rugosa n.gen.? n.sp. 1; LPB 17 254, thin
section a. B, longitudinal section
showing extrazooidal vesicular tissue and some section of lunaria which fibrous
microstructure is well exposed; LPB 17 254, thin section a. Monfoulon,
Armorican Massif, Monfoulon Limestones, Upper Pragian. |
The modalities of the association coral-bryozoan, involving for example Ordovician (Vinn et al., 2017a), Devonian (Sendino et al., 2019) or Neogene (Cadée & Mckinney, 1994) species, have been discussed and it appears that the true nature of the association is difficult to assess.
Cadée and Mckinney (1994) dealing with the intergrowth Celleporaria palmata - Culicia woodii discussed possible beneficial and negative effects for the coral and the bryozoan and concluded that "the association is a commensal or, more likely, a mutualistic relationship, probably obligatory for the coral, but non-obligatory for the bryozoan". These conclusions largely apply to our material but as to whether or not the association is obligatory for the coral remains uncertain.
Vinn et al. (2017a) stated that the so-called symbiotic association of Stigmatella massalis - Lambelasma sp. is "presumably purely accidental". It can be true, but in the Armorican and the Ougarta material - taking into account the rather numerous cases of the same kind of association - it seems that the corals selected most suitable organisms for their growth and that massive Cystoporata provided a convenient place.
An example of intergrowth between likely the same kind of coral and a Chaetetida from the Lower Emsian of the Middle Harz Mountains (locality Grosses Mühlental, South of altenbrak, coll. D. Weyer, LPB 17 259ter, not described herein) supports this idea. In this example the general morphology of the chaetetid is similar to that of the bryozoan and their small tubular units similar by their size to the zooecia of the bryozoan. The locality belongs to the Giessen-Harz Nappes and thus, probably to North Gondwana (Plusquellec & Jahnke, 1999).
On the other hand, as far as we know the symbiosis concerns two organisms physiologically connected and unable to live one without the other (Pouyet, 1978), thus the somewhat atypical example ("large rugosan partially embedded within the bryozoan colony") provided by Vinn et al. (2017a) belongs more likely to commensalism than to symbiosis.
The presence of rugosan - bryozoan intergrowth in the Lower Devonian of North Gondwana (Armorican Massif and Ougarta area) is documented and widely illustrated. This association is not unusual.
The Armorican association involves an unidentified Triplasmatidae? assigned herein to n.gen.? n.sp. 1 and some Cystoporata: n.gen. 1 cf. Crepipora, rarely Fistulipora sp. or to another unidentified Fistuliporidae.
The wall of the bryozoan zooecia is generally lacking where the bryozoan is in direct contact with the Rugosa.
The rugosan-bryozoan intergrowth can be followed by immuration of (very likely) dead corallites.
The intergrowth gives rise to a pseudo-colony of Rugosa.
The presence of rugosan-bryozoan intergrowth is not restricted to the Ordovician of Baltica and Laurentia being now well documented in the Lower Devonian of Gondwana (Armorican Massif, Cantabrian Mountains and Ougarta Mountains).
The association of Triplasmatidae-like corals with chaetetids is briefly indicated in the lower Emsian of Harz Mountains.
The association is very likely a mutualistic relationship if, as stated by Cadée and Mckinney, "the stinging cells of the coral provided protection against predator" and the bryozoan provided substrate and "the surrounding of the corallites by the bryozoan strength and lateral protection".
Except for the possible inclusion of the genus Ganiella (poorly known and family placement uncertain) and of two very doubtful Crepipora from Asia, the Ceramoporidae have not been indisputably recorded in the Devonian. The present paper provides an example of their presence in the Lower Devonian of the Armorican Massif. However, the Upper Ordovician Crepipora with lamellar microstructure are replaced in the Devonian by Crepipora-like forms with fibrous autozooecial wall, belonging consequently to a new genus.
The manuscript was critically read by O. Vinn, University of Tartu, Estonia and P. Fedorov, St. Petersburg State University, Russia. . The "LFP" thin sections were made by M. Lemoine of the Muséum National d'Histoire Naturelle, Paris. The photography of the thin sections was done by J.-P. Cuif in the same institution, the Photoshop © matters by B. Granier, Université de Bretagne Occidentale, Brest, and the linguistic review as well as some scientific suggestions by A.J. Wright, University of Wollongong, N.S.W., Australia. The authors are deeply indebted to the above persons for their help and assistance.
Boardman R.S., Cheetham A.H., Blake D.B., Utgaard J., Karklins O.L., Cook P.L., Sandberg P.A., Luteaud G. & Wood T.S. (1983).- Treatise on Invertebrate Paleontology, Part G Bryozoa Revised.- The geological Society of America, Boulder; The University of Kansas, Lawrence, 625 p.
Cadée G.C. & McKinney F.K. (1994).- A coral-bryozoan association from the Neogene of northwestern Europe.- Lethaia, Oslo, vol. 27, p. 59-66.
Coen-Aubert M., Gourvennec R., Monod O., Plusquellec Y. & Tourneur F. (2013).- Frasnian rugose and tabulate corals from the eastern Taurus (Kozan region, Turkey).- Geodinamica Acta, Zagreb, vol. 26, no. 1-2, p. 68-93.
Cuif J.-P. & Gautret P. (1993).- Microstructural features of fibrous tissues in the skeletons of some chaetetid sponges.- Courier Forschungsinstitut Senckenberg, Frankfurt am Main, vol. 164, p. 309-315.
Fedorowski J. (1991).- Principles of early ontogeny in the rugose corals: a critical review.- Hydrobiologia, Belgique, vol. 216/217, p. 413-418.
Hill D. (1981).- Treatise on Invertebrate Paleontology, Part F Coelenterata, Supplement 1 Rugosa and Tabulata, vol. 1.- The geological Society of America, Boulder; The University of Kansas, Lawrence, 378 p.
Kershaw S. (1987).- Stromatoporoid-coral intergrowths in a Silurian biostrome.- Lethaia, Oslo, vol. 20, p. 371-380.
Lafuste J. (1962).- Note préliminaire sur la microstructure de la muraille chez Favosites Lamarck (Coelenterata, Tabulata).- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, no. 4, p. 105-106.
Lafuste J. & Tourneur F. (1991).- Biocristaux et éléments foncés de la muraille chez Thamnopora Steiniger, 1831 (Tabulata, Dévonien).- Annales de Paléontologie, Paris, Vol. 77, fasc. 1, p. 3-20.
Le Menn J. (1985).- Les crinoïdes du Dévonien inférieur et moyen du Massif armoricain.- Mémoires de la Société géologique et minéralogique de Bretagne, Rennes, vol. 30, 268 p. (39 Pls.).
Michelin H. (1847).- Iconographie zoophytologique : Description par localités et terrains des polypiers fossiles de France et pays environnants.- P. Bertrand, Paris, 348 p. URL: https://www.biodiversitylibrary.org/item/43749#page/9/mode/1up
Milne-Edwards H. & Haime J. (1850).- A monograph of the British fossil corals.- Monographs Palaeontological Society London, vol.3, I-LXXXV, p. 1-72.
Modzalevskaja E.A. & Orlovsky M.B. (1968).- Novye vidy Siluriiskikh i Devonskikh mchanok srednei Asii. In: Markovskii B.V. (ed.), Novye vidy drevnikh rasteniĭ i bespozvonochnykh SSSR.- Akademia Nauk SSSR, Bashkirskii Filial Institut Geologii, Moskva, vol. 2, pt. 2, p. 47-54.
Paris F., Boumendjel K., Morzadec P. & Plusquellec Y. (1997).- Synthèse chronostratigraphique du Dévonien de l'Ougarta (Sahara occidental, Algérie).- Annales de la Société géologique du Nord, Lille, tome 5, p. 117-121.
Plusquellec Y. & Bigey F. (1990).- Une association tétracoralliaire-bryozoaire dans l'Emsien nord-gondwannien. In: Congrès national de Paléontologie, Paris 1990.- Société géologique de France, Paris, p. 87.
Plusquellec Y. & Jahnke H. (1999).- Les tabulés de l'Erbslochgrauwake (Emsien inférieur du Kellerwald) et le problème des affinités paléogéographiques de l'allochone ' Giessen-Harz'.- Abhandlungen der Geologischen Bundesanstalt, Wien, vol. 54, p. 435-451.
Plusquellec Y., Tourneur F. & Lafuste J. (1993).- Saouraepora nouveau genre de Micheliniidae (Tabulata), du Dévonien du nord Gondwana et du Carbonifère d'Amérique du Nord.- Palaeontographica A, Stuttgart, vol. 227, p. 1-86.
Pouyet S. (1978).- L'association bryozoaires - cnidaires.- Réunion annuelle des Sciences de la Terre, Paris, p. 319.
Semenoff-Tian-Chansky P. (1984).- Microstructure of Siphonodendron (Lithostrotionidae).- Palaeontographica Americana, Ithaca, vol. 54, p. 489-500.
Sendino C., Suárez Andrés J.L. & Wilson M.A. (2019).- A rugose coral - bryozoan association from the Lower Devonian of NW Spain.- Palæogeography, Palæoclimatology, Palæoecology, vol. 530, p. 271-280.
Sorauf J.E. & Kissling D.L. (2012).- Rugosans immured in Silurian Paleofavosites; Brassfield Formation (Llandovery) of Ohio.- Geologica Belgica, Bruxelles, vol. 15, no. 4, p. 220-225.
Spjelnaes N. (1963).- A redescription of the type species of the bryozoan genus Crepipora.- Journal of Paleontology, Menasha, vol. 37, no. 1, p. 64-68.
Stolarski J. (1996).- Gardinera - a scleractinian living fossil.- Acta Palaeontologica Polonica, Warszawa, vol. 41, no. 4, p. 339-367.
Vinn O., Ernst A. & Toom U. (2016).- Earliest symbiotic rugosans in cystoporata bryozoan Ceramopora intercellata Bassler, 1911 from Late Ordovician of Estonia (Baltica).- Palæogeography, Palæoclimatology, Palæoecology, vol. 461, p. 140-144.
Vinn O., Ernst A. & Toom U. (2017a).- Rare rugosan-bryozoan intergrowth from the Upper Ordovician of Estonia.- Carnets Geol., Madrid, vol. 17, no. 7, p. 1-6.
Vinn O., Liang K. & Toom U. (2017b).- Endobiotic rugose coral symbionts in Silurian tabulata corals from Estonia (Baltica).- Palaios, Lawrence, vol. 32, p. 158-165.
Vinn O., Toom U. & Ernst A. (2018).- Intergrowth of Orbignyella germana Bassler, 1911 (Bryozoa) and Lambelasma carinatum Weyer, 1993 (Rugosa) in the pelmatozoan-bryozoan-receptaculitid reefs from the Late Ordovician of Estonia. Palaeontologia Electronica, Article 21.1.4A, https://doi.org/10.26879/818, 4 p.
Vinn O., Wilson M.A., Toom U. & Mõtus M.-A. (2015).- Earliest known rugosan-stromatoporoid symbiosis from the Llandovery of Estonia (Baltica).- Palæogeography, Palæoclimatology, Palæoecology, vol. 431, p. 1-5.
Weyant M., Bultynck P., Plusquellec Y. & Racheboeuf P.R. (2010).- Reassessement of Lochkovian - Pragian conodont faunas from the Rade de Brest and the Presqu'île de Crozon (Massif Armoricain, France).- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, Abh. 258/1, p. 73-88.