![]()
Contents
[1. Introduction] [2. Description of the thuoux section proposed as the GSSP of Oxfordian stage]
[3. The Callovian-Oxfordian boundary and the fossil record]
[4. Satisfaction of geological requirements for Thuoux GSSP proposal]
[5. Correlations and comparisons with the alternative candidate GSSPs]
[Bibliographic references]
Le clos des Vignes, F-26160 La Begude de Mazenc (France)
8a, avenue Ste Claire, F-06100 Nice (France)
12, rue des Vergers, F-21121 Hauteville-les-Dijon (France)
29, rue Sainte Colombe, F-94800 Villejuif (France)
7, rue Albert Ier, F-94120 Fontenay-sous-Bois (France)
* Chercheurs libres au Centre des Sciences de la Terre, Université de Bourgogne, 6 boulevard Gabriel, 21000 Dijon (France)
Manuscript online since 14 July 2012
[Editor: Bruno ; copy editor: Christian C. ;
language editor: Stephen ]
![]()
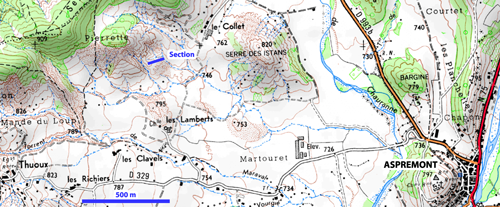
The Thuoux section, located in South-Eastern Basin of France (coordinates: 44°30'55"E; 5°42'25"N), is a section that satisfies numerous demanding criteria as reference section (GSSP) for the base of the Oxfordian stage. Sedimentation was continuous in that the abundant ammonitic fauna yields no detectable hiatuses. The stratigraphic boundary is located between the Lamberti Zone and the Mariae Zone or more precisely between the Paucicostatum horizon (, 1979) and the Thuouxensis horizon ( & , 1994a). In this section, there is a perfect mixing between Boreal ammonites (Cardioceratinae) whose species are used as stratigraphic markers and Submediterranean/Subboreal ammonites (Hecticoceratinae, Peltoceratinae and Perisphinctinae) that provide further possibilities for wide correlation of this boundary. Finally, parallel ammonite zonations have been established with great precision (biohorizons and sometimes "sub-biohorizons") in various areas of France, with different palaeoenvironments. The Thuoux section permits correlations with central and eastern Europe as well as North America, based on Cardioceratinae successions, and with South America, using Peltoceratinae. Thickness of the Callovian-Oxfordian transitional beds in the Thuoux section has allowed detailed sedimentological studies and astronomical calibration of the Lower Oxfordian (, 2008; et al., 2008). The Thuoux section is located at the centre of a set of more than thirty sections where the base of the Oxfordian stage is visible (, 1989; & , 1994a, b, c, d).
Ammonites; upper Callovian; lower Oxfordian; GSSP Oxfordian.
D., D., A., R. & (2012).- Proposal for the Thuoux section as a candidate for the GSSP of the base of the Oxfordian stage.- Carnets de Géologie [Notebooks on Geology], Brest, Article 2012/06 (CG2012_A06), p. 117-136.
Proposition de candidature de la coupe de Thuoux (France) pour le GSSP de la base de l'étage Oxfordien.- La coupe de Thuoux, localisée en France dans le Bassin du Sud-Est, (coordonnées : 44°30'55"E; 5°42'25"N) satisfait à la majorité des critères demandés pour devenir le GSSP de la base de l'étage Oxfordien. La sédimentation est continue à l'échelle de l'horizon ammonitique et en particulier des horizons à Paucicostatum (, 1979) et à Thuouxensis ( & , 1994a). Dans cette coupe, il y a un mélange constant entre les ammonites boréales (Cardioceratinae) et les ammonites subboréales et sub-méditérranéennes (Hecticoceratinae, Peltoceratinae et Perisphinctinae) ce qui autorise de larges corrélations géographiques. De plus, la présence simultanée de ces 4 sous-familles permet un découpage temporel très précis et permet aussi de proposer des corrélations temporelles entre l'Europe Occidentale, le nord de l'Amérique (grâce aux Cardioceratinae) et l'Amérique du Sud (grâce aux Peltoceratinae). La forte épaisseur des sédiments à Thuoux a aussi permis des études sédimentologiques et une calibration astronomique de l'Oxfordien inférieur (, 2008 ; et al., 2008). Enfin, plus d'une trentaine de coupes ont été analysées en détail autour de Thuoux, ce qui augmente la fiabilité des résultats obtenus sur la coupe oů la base de l'Oxfordien est visible (, 1989 ; & , 1994a, b, c, d).
Ammonites ; Callovien supérieur ; Oxfordien inférieur ; GSSP Oxfordien.
The Thuoux and Savournon sections were proposed as candidates for Global Boundary Stratotype Section and Point (GSSP) of the Oxfordian Stage more than 15 years ago ( & , 1994a, b, c, d). Both these sections were assessed as excellent insomuch as in 1996 (in ) a vote by Oxfordian Working Group yielded the following results:
Subsequently, we have focused our efforts on the Thuoux section which is located about 600m N-E from Thuoux (close to Aspremont and Saint-Pierre-d'Argençon), 5km
west of Aspres and 7km north of Serres (Département des Hautes-Alpes, France). Coordinates: 44°30'55"E; 5°42'25"N (Figs. 1 ![]() - 2
- 2 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 1: Location of the
Thuoux section proposed as GSSP Oxfordian.
© www.geoportail.fr
& IGN - Institut Géographique National, 73 avenue de Paris, F-94165 Saint-Mandé Cedex (France)
The Thuoux section, in contrast to Savournon, is not disturbed by faults and provides readier recognition of the units described below. Moreover, the gentler slope of the Thuoux section precludes strong mixing of faunas as well as pollution of the section by elements coming from the upper layers.

Click on thumbnail to enlarge the image.
Figure 2: The Thuoux section with the position of the Callovian / Oxfordian boundary (C: Callovian and O: Oxfordian).
(2004, 2006) expressed doubts about choosing Thuoux "because of preservation of ammonites as pyritic nuclei" or "the poor state of preservation of ammonites, as small pyritic nuclei, making them difficult to interpret". As a consequence, in the table p. 18 of (2006), ammonites from Redcliff are assessed (5) against Thuoux (1). This is surprising as Thuoux and Savournon ammonites are preserved in a virtually identical way ( & , 1994c, d). At both Thuoux and Savournon, the fossils are carbonate internal moulds corresponding either to the phragmocone alone, or including part or all of the body chamber. Never are the ammonites pyritic. In both sections, the average diameter of ammonites is about 5 cm.
In the South-Eastern Basin of France a very thick marly
limestone series bearing white and carbonate nodules crops out. The succession is known under the name of "Terres Noires", a term which also denotes its facies. This unit
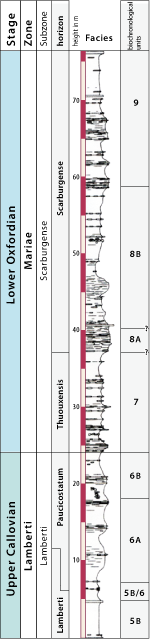
ranges from upper Bajocian to middle Oxfordian (Antecedens Subzone). At the Thuoux section, the stratigraphic interval from the upper Callovian (Lamberti Zone, Lamberti Subzone) to the lower Oxfordian (Mariae Zone, uppermost Praecordatum Subzone) is well
preserved and almost 200m in thickness (Fig. 3 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 3: Log (after Emilia ) and biostratigraphy (D. and D. ) of the "Thuoux" section.
This study was initiated by during the
1980s, resulting in 1989 in the first publication in which detailed subdivision of this section was established. That
biostratigraphical framework was later updated ( &
, 1994a; et al.,
1997) and officially accepted by the Groupe Français d'Étude du Jurassique
( et al., 1997). At present, it is accepted by
all Western European colleagues and notably by in Weymouth
(1996, 1997, 1999) and
(2010) in
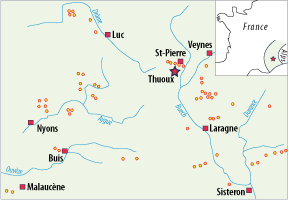
the French Jura. After more than 25 years' research, the Callovian/Oxfordian transition has been observed in the South-Eastern Basin
in 61 sections (Fig. 4 ![]() ), and in thirty of them exhaustive in situ
collection of ammonites has been carried out, in order to
determine precisely the limits of every horizon to analyse the ammonite populations (,
1989; et al.,
1990; et al.,
1997). More than 5,000 ammonites were collected close to the upper Callovian/lower Oxfordian boundary.
It is particularly important to note that in all these collections, from all levels, Phylloceratidae and especially Sowerbyceras, are always present and
abundant.
), and in thirty of them exhaustive in situ
collection of ammonites has been carried out, in order to
determine precisely the limits of every horizon to analyse the ammonite populations (,
1989; et al.,
1990; et al.,
1997). More than 5,000 ammonites were collected close to the upper Callovian/lower Oxfordian boundary.
It is particularly important to note that in all these collections, from all levels, Phylloceratidae and especially Sowerbyceras, are always present and
abundant.
Click on thumbnail to enlarge the image.
Figure 4: Locality map of the 61 outcrops in the "Terres Noires" Formation of the Southern-Eastern Basin of France where the Callovian-Oxfordian boundary is recognized.
At the levels 5B, 6A-6B, 7 and 8A (field numbers) which correspond to the Callovian/Oxfordian transition (, 1989), ammonitids are more abundant than phylloceratids, whereas from levels 8B/9 (uppermost Scarburgense biohorizon and lowermost Woodhamense biohorizon) we record the reverse. The latter is interpreted as due to the occurrence of extensional tectonics correlated with deepening and so with increase of phylloceratids. This phenomenon is known from numerous regions of Western Europe (, 1986).
Faunal comparisons were effected between the Boreal and Tethyan provinces, thanks to very precise collections made on the Isle of Skye (, 1963; , 1975; & , 1995) and around Nice ( & , 1979), and in between in Yorkshire, Dorset, the margins and centre of Paris Basin (Agence nationale pour la gestion des déchets radioactifs, ANDRA drillings), the Jura, the South-Eastern Basin of France, and several European countries (Germany, Bulgaria, Poland and Rumania with the help of colleagues in these countries).
Among all the Western European sections (see references), the Thuoux section is clearly the most accessible, the most interesting and the most reliable. It begins with upper Callovian (Lamberti Zone) and ends with lower Oxfordian (slightly below the Mariae Zone/Cordatum Zone boundary) deposits. This stratigraphic interval is continuous and, quite rarely for the "Terres Noires", free of faults. Moreover, sedimentary layers are sub-vertical, which makes thickness measurements easy. The occurrence of thin, slightly more carbonate-rich levels leads, through differential erosion, to smooth recesses between carbonate-rich intervals, which facilitates the collecting of ammonites in situ. There are also reddish carbonate intervals and concentrations of aligned grey to ochre nodules that constitute highly visible and very reliable markers. In these sections, ammonites are frequent, and commonly of greater size than in Savournon, with shell diameter above 10 cm. Counts made by D.F. and D.M on their collections from Thuoux are as follows ( & , 1994d):
Collected macro-invertebrates are low in diversity as ammonites frequently reach 100% of the macrofauna. Other cephalopods are present but belemnite rostra and rhyncholithes are scarce, and nautilids extremely rare. Bivalves are not frequent although sometimes they are concentrated inside ammonite body chambers (levels 7B and 9). Brachiopods and echinoderms are totally absent at Thuoux. And what is true at Thuoux is true within all the "Terres Noires" of the Dauphinois Basin.
a. Ammonites ( 1979; & , 1994a, b, c, d)
Uppermost Callovian ammonites (Lamberti Zone, Lamberti Subzone, Lamberti and Paucicostatum biohorizons).


Click on thumbnail to enlarge the image.
Plate 1: 1. Alligaticeras aff. alligatum () (M). Level 5B, Lamberti Zone, Lamberti Subzone, Lamberti horizon, n° DF 1741: Thuoux.
2. Poculisphinctes poculum () (M). Level 5B, Lamberti Zone, Lamberti Subzone, Lamberti horizon, n° DF 4588: Thuoux.
3. Quenstedtoceras lamberti () (M). Level 5B, Lamberti Zone, Lamberti Subzone, Lamberti horizon, n° DF 2584: Thuoux.
4. Kosmoceras duncani () in ,
1976 (fig. 6-2) (M). Level 5B, Lamberti Zone, Lamberti Subzone, Lamberti horizon, n° DF 18698: Thuoux.
5. Hecticoceras (Orbignyceras) paulowi (de ) (M). Level 5B, Lamberti Zone, Lamberti Subzone, Lamberti horizon, n° DF 2467: Thuoux.
6. Hecticoceras (Putealiceras) punctatum (m). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 1376: Thuoux.
7. Hecticoceras (Putealiceras) punctatum (M). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2503: Thuoux.
8. Cardioceras paucicostatum (M ?). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2469: Thuoux.
9. Hecticoceras (Lunuloceras) pseudopunctatum (M ?). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2482: Thuoux.
10. Poculisphinctes sp. aff. poculum () (m). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 18082: Thuoux.
11. Hecticoceras (Lunuloceras) pseudopunctatum (m). Level 6B, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2558: Thuoux.
12. Cardioceras aff. paucicostatum (m ?). Level 6B, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2530, Thuoux.
13. Peltoceratoides eugenii () (m). Level 6B, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 18098, Thuoux.
14. Peltoceratoides eugenii () (M). Level 6B, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2573: Thuoux.
All specimens are from D. 's collection. Photos: D. ;
preparation of plate: Simone . Red point: end of phragmocone.
Note that the genera Kosmoceras and Distichoceras, clearly present in the last horizon of the Lamberti Subzone (5B),
have never been collected in the Paucicostatum horizon (level 6) of Western
Europe. Hecticoceratins (about 50% of fauna) abound but still display "Callovian" morphology (Pl. 1 ![]() : 6, 7, 9, 11). A quite rare species, Hecticoceras coelatum , is characterized by ventral ribs that
are continuous around the ventral axis. In contrast, in the specimens of H. coelatum , from
the Lamberti biohorizon (5B), these ribs never cross the ventral axis. In every French record of this species, transition from one morphology to
the other quite
reliably indicates the Callovian / Oxfordian boundary.
: 6, 7, 9, 11). A quite rare species, Hecticoceras coelatum , is characterized by ventral ribs that
are continuous around the ventral axis. In contrast, in the specimens of H. coelatum , from
the Lamberti biohorizon (5B), these ribs never cross the ventral axis. In every French record of this species, transition from one morphology to
the other quite
reliably indicates the Callovian / Oxfordian boundary.
Within this biohorizon the history of the two important genera also diverges:
Cardioceratinae's history changes with the appearance of genus Cardioceras (C. paucicostatum ) whose successive species are used as biostratigraphic markers up to the end of the mid-Middle Oxfordian;
Peltoceratinae's history begins to change, slightly later, through the appearance of the first member of
Peltoceratoides with P. eugenii () (Pl. 1 ![]() : 13, 14), a species with split ventro-lateral tubercles. Successive species will also be of use
in the biostratigraphic subdivision of the Lower Oxfordian (,
1995; et al.,
1997, 2002). In the level 6A, Peltoceratoides
aff. shrœderi does not show split tubercles (Pl. 2
: 13, 14), a species with split ventro-lateral tubercles. Successive species will also be of use
in the biostratigraphic subdivision of the Lower Oxfordian (,
1995; et al.,
1997, 2002). In the level 6A, Peltoceratoides
aff. shrœderi does not show split tubercles (Pl. 2 ![]() : 1).
: 1).
The combination of these two genus lineages (Cardioceras and Peltoceratoides) permit a very precise and reliable biostratigraphy.
Basal Oxfordian ammonites (Mariae Zone, Scarburgense Subzone, Thuouxensis, Scarburgense and Woodhamense biohorizons) (Pls. 2 ![]() - 3
- 3 ![]() ).
).


Click on thumbnail to enlarge the image.
Plate 2: 1. Peltoceratoides aff. schrœderi () (M). Level 6A, Lamberti Zone, Lamberti Subzone, Paucicostatum horizon, n° DF 2655: Thuoux.
2. Cardioceras aff. paucicostatum ( & ) (M). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 1365: Thuoux.
3. Hecticoceras (Brightia) thuouxensis & (m). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 2605: Thuoux.
4. Hecticoceras (Brightia) thuouxensis & (M). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 2311: Thuoux.
5. Hecticoceras (Brightia) thuouxensis & (m). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 18156: Thuoux.
6. Euaspidoceras armatum (de ) (M). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 1457: Thuoux.
7 a, b. Peltoceratoides eugenii () morphe eugenii (M). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 1666: Thuoux.
8 a, b, c. Peltoceratoides eugenii () morphe eugenii (M). Level 7, Mariae Zone, Scarburgense Subzone, Thuouxensis horizon, n° DF 2655: Thuoux.
9 a, b, c. Hecticoceras coelatum (). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, n° DF 1617: Thuoux.
10. Cardioceras aff. scarburgense & (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, n° DF 18751: Thuoux.
11. Cardioceras aff. scarburgense & (m?). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, n° DF 18752:
Thuoux.
All specimens are from D. 's collection.
Photos: D. ; preparation of plate: Simone
. Magnification: black scale bar = 1 cm, except
fig. 9c: red scale bar = 1.5 cm. Red point: end of phragmocone.
"Les Richiers" section, ammonites are characterized by occurrence (up to 30% of fauna), of a distinctive Hecticoceratinae taxon: Hecticoceras (Brightia) thuouxensis et al. (1997) (Pl. 2
: 3, 4, 5). This index species is unknown below (in Paucicostatum horizon) and becomes rare at the base of the Scarburgense biohorizon.
The Thuouxensis biohorizon was first described in the South-Eastern Basin, where it was identified in more than 30 sections. Recently it was also described in the Paris Basin from ANDRA drillings ( et al., 2006). Throughout the Thuouxensis biohorizon, the bifurcation points of Peltoceratoides always lie in the upper half of the lateral flank ( et al., 1997, 2002) (Pl. 2
This level corresponds to the Scarburgense biohorizon. The index species is easy to recognize (Pl. 2
In the sub-Tethyan Province, this Scarburgense biohorizon is also characterized by Hecticoceras with less pronounced sculpture such as H. chatillonense (Pl. 3
Peltoceratins from level 8B (Pl. 3


Click on thumbnail to enlarge the image.
Plate 3: 1 a, b. Cardioceras aff. scarburgense & (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 3880: Thuoux.
2. Cardioceras aff. scarburgense & (m). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 3883: Thuoux.
3 a, b. Cardioceras scarburgense & (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 3881: Thuoux.
4. Hecticoceras (Brightia) sp. (m). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 1608: Thuoux.
5. Cardioceras scarburgense & (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 18419: Thuoux.
6. Hecticoceras (Brightia) cf. thuouxensis & (M?). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 18493: Thuoux.
7. Hecticoceras (Brightia) chatillonense de (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 18767: Thuoux
8. Peltoceratoides eugenii () morphe eugenii (M). Level 8A, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, lower part, n° DF 3717: Thuoux.
9. Peltoceratoides athletoides () morphe athletoides (M). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 1944: Thuoux.
10. Peltoceratoides athletoides () morphe athletoides (M). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 18563: Thuoux.
11. Hecticoceras (Brightia) chatillonense de (M). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 1875: Thuoux.
12. Eochetoceras villersensis (d') (m). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 1385: Thuoux.
13. Cardioceras scarburgense ( & ) (M?). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 1659: Thuoux.
14 a, b. Taramelliceras episcopalis (de ). Level 8B, Mariae Zone, Scarburgense Subzone, Scarburgense horizon, middle part, n° DF 19536:
Thuoux.
All specimens are from D. 's collection.
Photos: D. ; preparation of plate: Simone
. Red point: end of
phragmocone.
At Thuoux, this level corresponds to the Scarburgense biohorizon (upper part).
Abruptly, ammonites are less frequent and Cardioceras is very scarce. Records of Eochetoceras villersensis (d') and E. hersilia (d') at the base of that level,
as in the French Jura (, 2010), and the disappearance of Hecticoceras coelatum are typical of that level. From this stratigraphic level, Phylloceratina become more abundant than Ammonitina,
due to a well-known deepening in numerous sections in Western Europe ( et al.,
1994). In other sections, very similar to that investigated at Thuoux, the
top of this stratigraphic level has yielded some Cardioceras close to the species
C. woodhamense ( & ,
1994a; et al.,
nearing completion). C. woodhamense definitively appears above the Scarburgense horizon and not below, as
claimed by (1993) and et al.
(2006, 2009a, b).
Though ammonites are very rare, faunal assemblages indicate the Woodhamense horizon and the upper part of the basal Praecordatum Subzone.
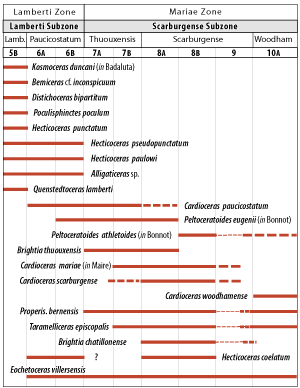
In conclusion, the correlation potential of ammonites collected from the Thuoux section is extremely precise (Fig. 5 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 5: Vertical range of the major species of ammonites found in the "Terres Noires" Formation at the Callovian-Oxfordian boundary (NB: 5B to 10A are field subdivisions).
In this paper, subdivisions proposed for the Thuoux GSSP are the same proposed in 1997 by et al. for the Callovian and et al. for the Oxfordian. They are known in all Western Europe and have been used without difficulty in the United Kingdom, Germany, Switzerland and Poland ( & , 1990), and only on the basis of Peltoceratins in Bulgaria and Rumania where Cardioceratins are unknown. Correlations may be also possible:
b. Foraminifera
On certain samples, (in , 1996) wrote: "Thuoux foraminifers and nannoplankton in the studied interval were low in diversity but moderate in abundance. Preservation was, in general, poor. Only two species recorded were stratigraphically significant". These two species are Ophthalmidium compressum and O strumosum (); O. compressum disappears at the Thuouxensis biohorizon where it is replaced by O. strumosum, the latter appearing in the Paucicostatum biohorizon and persisting into the Scarburgense biohorizon.
c. Dinoflagellates
This study was carried out by N.E. from sampling performed in 1994 during the 4th Oxfordian and Kimmeridgian Working Group Meeting. Results can be summarized as follows. "Most of the stratigraphical important species are present in the assemblages" (citations from in , 1996). Comparison of the range charts for the Savournon and Thuoux sections shows that important species occur in both sections. Moreover, (in , 1996) confirms that Durotrigia filapicata disappears at the top of the Callovian whereas Wanaea fimbriata appears at the base of the Oxfordian, but is very rare. These results are in accordance with what is known "in North Sea Region and in East Greenland". (in , 1996) also noted that "the preservation of the palynomorphs in the Thuoux section is poor and the number of cysts is low" and that "in the Savournon section dinoflagellate cysts are excellently preserved and present in large numbers".
d. Ostracodes
Preliminary research shows they seem to be rare but present at Savournon (, 2008).
e. Isotope stratigraphy
This may be feasible from rare belemnite rostra.
f. Magnetostratigraphy
All samples are negatively remagnetized (B. , oral communication).
The Thuoux section fulfills numerous requirements proposed in the Guidelines of the ICS, i.e., International Commission on Stratigraphy ( et al., 1996).
a. Geological requirements
Exposure over an adequate thickness (Fig. 2 ![]() ).
).
Biohorizons: Lamberti (5B), Paucicostatum (6A and 6B), Thuouxensis (7A and 7B) and Scarburgense (8A and 8B) reach 60m in thickness. Vegetation is very sparse and exposed levels are easy to recognize.
Continuous sedimentation (Fig. 3 ![]() ):
the faunal succession, together with the thickness of sedimentary rock, indicates very continuous
sedimentation.
):
the faunal succession, together with the thickness of sedimentary rock, indicates very continuous
sedimentation.
The rate of sedimentation: for the investigated stratigraphic interval in Western Europe, such a substantial thickness is known only in the "Terres Noires" area of the South-Eastern Basin (together with the centre of the Paris Basin).
NB: On the basis of biostratigraphic data, the sedimentation rate is 6 to 20 times higher than in Weymouth, UK.
Absence of synsedimentary and tectonic disturbances: the Thuoux section is undisturbed, and neither faults nor landslides have been noticed. Moreover, strong layer dip makes mixing of faunas unlikely.
Absence of metamorphism and strong diagenetic alteration: both phenomena are absent. Superficial erosion and weathering maintains the section in a very good state, which makes collection of samples easy for every type of geological study.
b. Biostratigraphic requirements
Abundance and diversity of well-preserved fossils: ammonites are abundant and well-preserved, and always greatly exceed other macroinvertebrates in abundance. They are often extracted, entire rather than crushed, from calcareous nodules. Fossils at Thuoux are characterised by a mineralogy that is stable in a weathering environment. Though (2003, 2004, 2006) asserted the occurrence of pyritic replacement of ammonites at Thuoux, any pyritic ammonites would have been altered to goethite. In fact, neither pyritic nor goethitic preservation is present! Boreal as well as Mesogean faunas can be collected. Moreover, the analysis of ammonite faunas in the South-Eastern Basin in the context of synsedimentary tectonics ( et al., 1994) has allowed better understanding of faunal distribution in relation to palaeodepth in Western Europe ( & , 1979; , 1984; et al., 1990). Microfossils are quite numerous and some groups can be used for some biostratigraphic schemes whose precision is, at its best, of subzonal scale.
Absence of vertical facies changes (Fig. 2 ![]() ):
the Callovian / Oxfordian boundary occurs in a series of marls that lack facies changes other than some fluctuations in the carbonate content.
):
the Callovian / Oxfordian boundary occurs in a series of marls that lack facies changes other than some fluctuations in the carbonate content.
Favourable facies for long-range biostratigraphic correlation (Fig. 3 ![]() ):
the Thuoux section yields three ammonite subfamilies (Cardioceratinae, Peltoceratinae and Hecticoceratinae) whose morphological evolution
was rapid, though not synchronous. These factors
ensure 1) the reliability of the faunal successions at the Callovian / Oxfordian boundary
and 2) the biostratigraphic precision of biostratigraphic units. These three
subfamilies also allow correlations not only with all Western Europe but also with
the Boreal Realm (North America) and the Pacific Realm (Argentina, Chile). As taxa of these three
subfamilies are not always found together, it is important to be able to use three parallel
schemes for biostratigraphic control.
):
the Thuoux section yields three ammonite subfamilies (Cardioceratinae, Peltoceratinae and Hecticoceratinae) whose morphological evolution
was rapid, though not synchronous. These factors
ensure 1) the reliability of the faunal successions at the Callovian / Oxfordian boundary
and 2) the biostratigraphic precision of biostratigraphic units. These three
subfamilies also allow correlations not only with all Western Europe but also with
the Boreal Realm (North America) and the Pacific Realm (Argentina, Chile). As taxa of these three
subfamilies are not always found together, it is important to be able to use three parallel
schemes for biostratigraphic control.
c. Other methods
Radioisotopic dating: not applied.
Magnetostratigraphy: numerous analyses were always negative in the South-Eastern Basin, in contrast to Redcliff Point.
d. Faunal populations
Ammonites: at each stratigraphic level examined, ammonites were collected in situ and exhaustively. Thus faunal diagrams
depict the faunal composition of each level (each biohorizon commonly being subdivided into two ammonitic
sections: lower / upper). They also summarize our knowledge of the
palaeodepth of the fossiliferous deposits studied. One of us has even used these faunal diagrams to establish
the occurrence of extensional tectonic events during the earliest Oxfordian (Scarburgense Subzone: ,
1986) and more precisely between the Scarburgense and Woodhamense biohorizons (,
1986) (Fig. 5 ![]() ).
).
Foraminifera: diversity is low, but the occurrence of Ophthalmidium compressum in the uppermost Callovian and Ophthalmidium strumosum () in the lowermost Oxfordian can be used for biostratigraphy.
Dinoflagellates: Durotrigia filapicata is rare in the uppermost Callovian (Lamberti horizon) and disappears in the lowermost Oxfordian (Scarburgense horizon) where Wanaea fimbriata , also rare, first occurs.
e. Other requirements
Permanently fixed marker: conservation of the section proposed as GSSP is assured at Thuoux. The Thuoux section is located in a huge area of badlands that are neither cultivable nor suitable for building.
Accessibility (Fig. 1 ![]() ):
access will be free for every researcher. Access is possible from a parking zone
(at the nearby cemetery) located 10 minutes' walk from the section, on a
gentle slope (30 m difference in height). Researchers will be able to work without
constraint by using the
same biostratigraphic subdivisions proposed by ammonite specialists (D.F. and
D.M). Permanent marking of the biostratigraphic units recorded by the authors is easy to make.
):
access will be free for every researcher. Access is possible from a parking zone
(at the nearby cemetery) located 10 minutes' walk from the section, on a
gentle slope (30 m difference in height). Researchers will be able to work without
constraint by using the
same biostratigraphic subdivisions proposed by ammonite specialists (D.F. and
D.M). Permanent marking of the biostratigraphic units recorded by the authors is easy to make.
a. Redcliff Point, Weymouth, Dorset (UK)
This section, well analysed by et al. (2006, 2009a), has six shortcomings, here addressed in order of decreasing importance.
* The first disadvantage is clearly acknowledged by the British authors themselves: the section is not reliably exposed. "For some time, the exposure (Redcliff Point) has been covered by talus and vegetation, but recent coastal erosion has now re-exposed a Callovian-Oxfordian boundary sequence" (in , 1994a, b). "The extent and quality of the exposure at Ham Cliff varies from season to season and from year to year, depending on the height of the shingle storm beach and cliff erosion" (in , 2003: 30, p. 17). "The section has been obscured by slumping in recent years" (in , 2004: 31, p. 9). "Correlation between these descriptions by different authors has been difficult due to the variable condition of the exposure and the non persistence of various beds taken as marker horizons" (in et al., 2006). "The soft nature of the mudrock outcrop makes the placing of a permanent marker problematic" (in et al., 2009b, p. 107). But et al. (2009b, p. 107) wrote in conclusion that "although the area is subject to contemporary landslipping, continuous cross-boundary sequences remain available."
We therefore spent many hours at this section with in 2003 and we ( & ) confirm that it is very difficult, even for Callovian / Oxfordian boundary specialists, to recognise the lithological succession described by et al. (2009a, b: fig. 2, p. 105). (in & , 1993) had said about the Ham Cliff section -close to Redcliff Point- (quoted by G. : 2003) "The reason why it had been neglected since 's days was that it was usually obscured by slumping and shingle. I remember looking at it in 1956 and actually finding the Lamberti Zone there, in a small outcrop above the shingle. But all I learned was that it confirmed 's report of an occurrence which had been ignored previously. Exceptional storm conditions in the winter of 1991-1992 swept it all clear, giving a fine exposure". And (in & , 1993) described an exceptionally clean section at Redcliff Point where C. woodhamense occurs below C. scarburgense ( & ). Unfortunately, et al. (2009a) have accepted that mistaken succession.
* The second drawback is palaeontological: the faunal assemblage is poorly preserved, with rather low diversity. The assemblage has not been illustrated, except for the Cardioceratins (1 plate). It should be emphasized that Cardioceratins predominate at Redcliff Point, but are "typically crushed" (though not always the inner whorls). This makes observation of the venter very difficult so that no fully reliable determination is possible. In addition, Hecticoceratinae are rare and the absence (in et al., 2009a) of Hecticoceras (Brightia) thuouxensis & does not allow recognition of the Thuouxensis biohorizon. However, this species is present close to Redcliff Point: it has been figured by (1999: Pl. 7, fig. F) in association with H. (B.) chatillonense de ( 1999: Pl. 7, figs. G-H). H. (B.) thuouxensis was also collected in Paris Basin boreholes ( et al., 2006) and in the Jura (, 2010). It is not a rare species at that level. Finally, individuals belonging to the genus Peltoceratoides, which is present though not abundant, were not used by and collaborators as biostratigraphic markers even though they are chronologically as precise as Cardioceras at the Callovian / Oxfordian boundary in the South-Eastern Basin of France ( et al., 1997; et al., 1997, 2002).
* The third disadvantage concerns ammonite systematics. The et al. (2009a, b) proposal relies on a dubious determination of Cardioceratinae species as well as an erroneous phylogeny.
Cardioceras paucicostatum is close to C. scarburgense ( & and must be included in the genus Cardioceras. Why is this? The first representatives of C. scarburgense ( & ) are in morphological continuity with paucicostatum , which is confirmed by the existence of "features transitional to C. (Pavloviceras) scarburgense ( & )". Both forms are present in horizons LL3c-d and Ox1 of et al. (2009a, p. 93) at Redcliff Point;
Cardioceras redcliffense et al. (2006), named Cardioceras (Pavloviceras) redcliffense (2009a), in our opinion has no palaeontological reality. We regard C. redcliffense et al. as a synonym of Cardioceras paucicostatum , 1973. As a matter of fact, in the Ox1 biohorizon, the holotype of C. redcliffense ( et al., 2009a: pl. 1 D) shows a C. scarburgense-like morphology but among a population where "paucicostatum" morphology is still abundant ( et al., 2009a: Pl. 1 C, E-G). Unfortunately, no information on Hecticoceratins and Peltoceratins is available to permit precise correlation with the Thuoux section.
Cardioceras scarburgense ( & ) from Scarburgense biohorizon is quite easy to differentiate from C. paucicostatum . The holotype of C. scarburgense, from Scarborough, was figured by (1939: Pl. X, fig. 1a-c). It is characterized by a) a moderate number of primary ribs, b) secondary ribs that are more or less prorsiradiate and c) a venter lacking crenulation in lateral view. These morphological characteristics are exactly the same:
- at Authoison (Département de Haute-Saône, France), where a large population of C. scarburgense shows exactly the morphological variations of that species, well illustrated by in 1938 (Scarburgense biohorizon);
- in populations of C. scarburgense collected in the levels 8A/8B and basal 9 from "Terres Noires" of French South-Eastern Basin ( & , 1994a);
- in populations of C. scarburgense collected at Uzelot (in Boulonnais) from the levels IT 20/30 and IT 30/40 ( et al., 1993);
- in the horizons named Scarbu (1) and Scarbu (2) from French Jura (, 2010).
Cardioceras woodhamense is for us clearly younger than C. scarburgense ( & ).
Remark: et al. (2009a: p. 92 and 95)
placed the Redcliffense biohorizon just below the Woodhamense biohorizon (Ox2) and accepted
the Scarburgense biohorizon (Ox3) above the Woodhamense biohorizon. Quite
evidently, that stratigraphy cannot be right.
In a forthcoming paper ( et al.,
nearing completion) we confirm through very close analysis of facts determined
from numerous sections that C. woodhamense is
younger than C. scarburgense ( & )
and that ammonite populations from the Scarburgense and Woodhamense horizons are clearly very different from each other.
Some insights from our results:
(i) About scarburgense ( & ) and woodhamense ( & , 1994b; et al., nearing completion).
- C. scarburgense ( & ) does not yet have an individualized keel, and shows no proverse ribs making a chevron (V-shape) on the venter;
- In contrast, the species C. woodhamense, erected by in 1939, incorporates an apomorphy that characterizes every later Cardioceras species: the presence of a keel or, more precisely, of primary and secondary ribs elevated on the ventral axis, forming a fairly prominent chevron. In lateral view, C. woodhamense shows a rather crenulated venter, which never occurs in C. scarburgense.
(ii) About Woodham Brick Pit Cardioceratinae and about C. redcliffense ( et al., nearing completion).
- Cardioceratinae from the Woodham Brick Pit, pictured by under the following names: Q. mariae, Q. omphaloides, Q. aff. williamsoni (all with undulating ribs in inner whorls like the holotype of C. woodhamense ), correspond to the "thick" and "medium thick" morphotypes of C. woodhamense .
- As consequence: the 157 C. scarburgense not pictured by are simply, from a biological point of view, the "thin morph" of C. woodhamense .
So our conclusion is that, at Woodham Brick Pit, there is a lacuna in place of the Scarburgense biohorizon because the Woodhamense biohorizon (levels A and B of , 1939) lies directly on the Upper Callovian (stratigraphic level C, Lamberti biohorizon, , 1939).
Remark: The same lacuna at the Scarburgense biohorizon is also known in Normandy (, oral communication).In summary, Cardioceras redcliffense ( et al., 2009a) looks like C. paucicostatum except for some individuals which are similar to C. scarburgense ( & ), such as the holotype ( et al., 2009a: Pl. 1D). No variants are close to C. woodhamense .
(iii) About the succession of ammonite assemblages in Western Europe (, 1986; et al., nearing completion).
(1939) exhaustively collected 899 ammonites from levels A and B at Woodham Brick Pit. The collection consists of Cardioceratinae (n = 297; 33% of the total assemblage), Hecticoceratinae (N = 102; 11.3 %), Perisphinctinae (n = 73; 8.1 %), Euaspidoceratinae (n = 44; 4.9 %) and also g. Taramelliceras (n= 281; 31.2 %) and Creniceras (n = 102; 11.3%).
In the French Jura (, 2010), as well as in the Swiss Jura (, 1990), Taramelliceras and Creniceras are always scarce in association with C. scarburgense, but always abundant with C. woodhamense, collected above. A similar succession occurs in Weymouth at Redcliff Point. (1999: Pl. 9) has pictured individuals that belong to C. woodhamense in particular, the individual of his fig. U, which is very similar to 's holotype. These C. woodhamense are associated with Taramelliceras sp. and C. renggeri, as at Woodham Brick Pit. These results are very clearly synthesized by (1999) in his fig. 10, p. 95.
In conclusion, the succession of biohorizons at Redcliff Point is actually: C. paucicostatum -> C. scarburgense -> C. woodhamense, instead of C. paucicostatum -> C. redcliffense -> C. woodhamense -> C. scarburgense.
* The fourth drawback is connected with sedimentation.
At Redcliff point, clays representing the youngest Callovian biohorizon and the oldest Oxfordian biohorizon are no thicker than 50 cm ( et al., 2009a: fig. 1). Thus, the following sentence is surprising: "Although the biohorizons are thinner, the Redcliff Point sections currently offer the potential for higher resolution than has been described for SE France" ( et al., 2009a).
It is particularly surprising since, at Thuoux, the thickness of the same horizons (levels 6B and 7) reaches about 20m, which is 40 times thicker than at Redcliff. However, the three palaeontologists that have worked at Thuoux were unable to identify within those 20 m more than two geographically widespread biohorizons, although the ammonite assemblages there are more diverse.
* The fifth drawback concerns the thickness of the section.
The small thickness observed at Redcliff Point makes any cyclostratigraphic analysis dubious. By contrast, such an analysis may be made in twenty sections of the South-Eastern Basin where precise biostratigraphic control is available. The Thuoux and Saint-Pierre-d'Argençon sections, for instance, could be sampled every 8 cm.
* The sixth drawback concerns correlations with the Boreal Domain and the Pacific Province.
In the Redcliff Point GSSP proposal (p. 90 - 91), et al. (2009a) have written about the "bioprovincial context" in general, without any attempt to correlate the various biostratigraphic schemes with each other.
For the Arctic Province, they write only that "there is no reason to suspect that the basic succession of Quenstedtoceras to Cardioceras faunas differs in any significant way from that in adjacent Subboreal areas". But they also claim that "there is some evidence of endemic cardioceratid in … North America".
For the Subboreal Province, they refer only to the domination of the Cardioceratids. On the Isle of Skye (Scotland), only Cardioceratids occur, except for one level which yields only Peltoceratoides sp. (, 1966). That stratigraphic level - very easy to find at Staffin Bay - marks the topmost Praecordatum Subzone, just a little below the Bukowskii Subzone ( & , 1979).
No confident conclusions can be drawn from the Russian forms, which are very well described by et al. (2004, 2005, 2006), nor from the South American and Pacific provinces.
For us, this is problematic because every GSSP must be, by definition, a reference point for any available biostratigraphic schemes. That is the true aim of a GSSP.
* Appendix: The case of Peyrale (Savournon, near Serres, Hautes-Alpes) proposed as GSSP auxiliary section.
Since 2003, has considered that the Peyrale section located close to Savournon "displays some particular features making it a suitable candidate section for Oxfordian GSSP". In 2007, in a new paper by , and (2007: fig. 3), these authors proposed this section as an "auxiliary section" of the Redcliff Point GSSP. The first 10m are attributed to the "Lamberti biozone" (levels 10 to 21) and the last 4-5 m (levels 19-21) yielded C. paucicostatum (= Q. paucicostatum in et al., 2009a). As in the Redcliff section, , and recognized three levels in the Paucicostatum horizon, which are from lower to upper:
C. paucicostatum-alpha "involute forms showing thick, bundled ribbing with low point of furcation";
C. paucicostatum-beta "more typical, thickly ribbed (non-bundled) evolute forms";
C. paucicostatum-gamma, "showing finer, slightly projected ribbing and the beginning of a ventral keel on innermost whorl"; these gamma forms are "common at level 21-c and become rare at bed 23a".
These authors place the "base of the Oxfordian stage at the base of the level 23 coinciding with the first recording of some specimens of Cardioceras of the group C. redcliffense et al., a new form displaying clear intermediate features between C. paucicostatum-gamma and C. scarburgense ( & )".
At this juncture, two points should be made. In our study of the Peyrale/Savournon section ( & , 1994d) we were able to state that:
Final remark: the Woodhamense biohorizon, which at Redcliff Point ( et al., 2009a) lies between the Redcliffense and Scarburgense biohorizons, vanishes in the Peyrale section. Since we know the Peyrale / Savournon section very well ( & , 1994b), we are aware that the Woodhamense biohorizon is present at Savournon. However we collected that species more than 25 m above the Paucicostatum horizon. As everywhere else, the Woodhamense biohorizon lies above the Scarburgense horizon, and not below.
b. Dubki section: Russia
Interesting information on the Callovian / Oxfordian boundary comes from our Russian colleagues and a short summary follows. According to and (2004, 2005) and et al. (2006), the Callovian / Oxfordian boundary is readily identifiable at Dubki, Saratov (52nd parallel). Thus, in 2006, they proposed the Dubki section as a candidate GSSP for the Lower Oxfordian boundary. We wish to address certain points related to this proposal. The Lamberti Zone (Upper Callovian), 4.60m thick, is subdivided into 2 subzones (Henrici and Lamberti) and 5 biohorizons (Henrici, Praelamberti, Lamberti, Mojarowskii and Paucicostatum).
The Mariae Zone, 2.60m thick, is subdivided into two subzones (Scarburgense, Praecordatum) and 3 biohorizons (Scarburgense, Alphacordatum and Praecordatum). In Russia, the genus Taramelliceras and the species Creniceras renggeri occur quite late (Bukowskii Subzone). Latest Kosmoceras disappears at the top of the Mojarowskii biohorizon (with the species K. mojarowskii), whereas the last occurrence of the genus Quenstedtoceras is registered slightly higher, at the Callovian / Oxfordian boundary, consistent with Western Europe.
Concerning ostracods, the species Infacythere dulcis disappears at the Callovian / Oxfordian boundary whereas Nophrecythere oxfordiana first occurs slightly above that boundary (, 2008). The same level is readily recognized from foraminifera which are represented by diverse and abundant benthic taxa. The Callovian / Oxfordian boundary coincides with the boundary between the Lenticulina tunida and Epistomina elschankensis zones and the Ophthalmidium sagittum and Epistomina volgensis zones. This section has yielded good palaeomagnetic results based on reversals, except at the Callovian / Oxfordian boundary.
In 2004, and had proposed an identical zonation for the Lamberti Subzone. In contrast, their proposed subdivision of the Mariae Zone differs slightly from our proposal.
At the base is the Scarburgense biohorizon from which they record two species, Vertumniceras luppovi () and Sublunoceras deperditum (), that are absent from assemblages in France, the United Kingdom and Switzerland. However, they have accepted the Woodhamense biohorizon as proposed by and in 1994b. This is reasonable as we regard the individual of Pl. 2: 17 of the former authors (Orenbourg area: in & , 2004) as being very close to C. woodhamense var. normandiana. and (2004) consider that the Praecordatum Subzone begins with an unnamed horizon that yields Cardioceras praecordatum and Peltoceras sp., overlain by the Renggeri horizon that yields Taramelliceras (Richeiceras), Creniceras, Peltoceras sp. and Cardioceras cf. alphacordatum. Their pictures of Q. henrici, Q. praelamberti, Q. lamberti, C. scarburgense, C. praemartini, C. alphacordatum, C. praecordatum and C. bukowski seem to us to be correctly identified except for C. costicardia (their fig. 11/12), C. cordatum (their fig. 13/14) and C. paucicostatum (their fig. 15/16). Species found in France and the United Kingdom have been shown to be present in Russia also, which makes correlation possible, as (1975) had already demonstrated.
The importance of the Dubki section is thus well established, in particular, due to mixing of "Boreal" and "Tethyan" faunas, which make long-distance correlations possible. Noticeable is that this faunal mixing, as frequently in Western Europe, occurs near the boundary between the Scarburgense and Praecordatum biohorizons whenever the palaeoenvironment was sufficiently open.
Nevertheless, the stratigraphic ranges of some species are not in strict accordance with what we know in the "Terres Noires" and at Thuoux in particular. For example the Paucicostatum biohorizon at Dubki yields both Poculisphinctes and Alligaticeras ( et al., 2002), and Euaspidoceras subbabeanum () sensu even crosses the Callovian / Oxfordian boundary;
Hecticoceratinae appears quite rare in that area and absent above the lowermost Oxfordian levels. Thus, general correlation with the Thuoux section is not yet demonstrated with that subfamily;
Although the faunal record shows considerable similarity with the Tethyan regions, there is no quantitative study yet.
It is important to note that some species recorded at Dubki have been identified from other deposits by and (2003) and and (2005). We are in agreement with almost all the proposed names. Accurate correlation with Western Europe is now possible with the help of these excellent papers.
c. Our proposals for correlation outside of Europe
* Argentina (biochronological interpretation: A. )
In "Biostratigraphy of the transect Chacay Melehue-Sierra de Reyes, Argentina", (1998) considered the dimorph pair Peltoceras gr. constantii (M) / arduennense (m) typical of the Cordatum Zone (lower part). The proposed occurrence of Prososphinctes gr. mazuricus / claramontanus and Prososphinctes gr. mairei / matheyi confirms that interpretation. By contrast Mirosphinctes gr. syriacus / regularis and Euaspidoceras cf. kobyi would rather suggest the Mariae Zone, and some individuals collected at Rahueco are similar to those described from Mount Hermon (Syria) for the lowermost Oxfordian. In our opinion, many specimens from Syria are very close to Eochetoceras villersensis / hersilia, a dimorph species that characterizes the top of the Lamberti Subzone up to the base of the Woodhamense biohorizon ( et al., nearing completion) in France and the United Kingdom.
Later, in 2006, compared ammonite successions of the Middle Callovian to Upper Oxfordian in both Neuquén-Mendoza and Tarapaca basins. Near to the Callovian-Oxfordian boundary he recognized:
Athleta Zone, demonstrated by the occurrence of Pseudopeltoceras sp. and Peltoceras (P.) cf. athleta;
Primus Zone that yields the genus Distichoceras (top of Athleta Zone / probable Lamberti Zone);
In the Neuquém-Mendoza Basin, the lower Oxfordian is represented by the Pressulus Zone only, which yields 3 species of genus Peltoceratoides, P. pressulus, P. ballenaensis and P. cf. intercissus. By comparison with Western European species, these species indicate the Scarburgense Subzone ( et al., 2002). However, if the identifications of Perisphinctes cf. mazuricus / matheyi are correct, this instead suggests the base of the Praecordatum Subzone;
In the Tarapacá Basin, the lower Oxfordian is represented by 3 zones; it begins with Peltoceratoides retrocostatum and P. dimorphosus (Dimorphosus Zone) and then continues with Peltoceratoides eugenii and P. ballenaensis (Eugenii Zone).
* Chile (biochronological interpretation: A. ).
and (1995) have described a biostratigraphic succession based on different species of the genus Peltoceratoides. They considered the Primus Zone to be equivalent to the Lamberti Zone, and Dimorphus and Eugenii zones equivalent to the Mariae and Cordatum zones.
As has been pointed out by et al. (2002), "if we roll out the hypothesis of pronounced endemism … the Chilean forms described … can all be ascribed either to Peltoceratoides eugenii (d') or to Peltoceratoides athletoides ()". As a consequence: their "Eugenii Zone" must be correlated with the Athletoides Subzone of the Mediterranean province = Scarburgense Subzone of Western Europe.
* North America (biochronological proposition: D. & D. ).
(1919) figured ammonites from Wyoming (Sundance Formation) and from Montana (Ellis Formation). In these regions, he recognized the genus Quenstedtoceras and 5 species. In our opinion, only Q. collieri (from the Ellis Formation) can be a true Questedtoceras. The other species (of the Sundance Formation) seem to have a small keel and are, probably, true Cardioceras. (1919) distinguished three groups of cardioceratids, for us, from the Mariae Zone:
* "Pacific area part" ( et al., 2002)
Peltoceratoides williamsoni () and allied species are found almost worldwide, and correlations can be envisaged between Western Europe and the Indo-Madagascar domain + Indonesia + Japan, where the genus Peltoceratoides is known.
We acknowledge the valuable comments on the initial draft by Raymond , the careful linguistic improvement provided by the Language Editor, Stephen , and the detailed remarks of the Referees of this paper, Nicol , Federico and Mikhail , who contributed to improve the scientific impact of this publication. Last but not least our thanks also go to the Copy Editor, Christian C. , and the Editor, Bruno .
W.J. (1939).- The ammonite succession at the Woodham Brick Company's Pit, Akenan Street Station, Buckinghamshire, and its bearing on the classification of the Oxford Clay.- Quarterly Journal of the Geological Society of London, vol. 95, p. 135-222.
A. (1976).- Biostratigraphical precisions upon the Middle Jurassic series in Anina (zone Resita, Banat).- Dări de seamă ale şedinţelor Institutului Geologic Bucuresti, 62, p. 73-84.
A. (1995).- Les Aspidoceratidae d'Europe occidentale au Callovien supérieur et à l'Oxfordien inférieur.- Thèse de Doctorat, Université de Bourgogne, Dijon, 487 p.
A., D. & D. (1997).- Les Peltoceratinae (Ammonitina, Aspidoceratidae) au passage Callovo-Oxfordien dans les "Terres Noires" du Sud-Est (France).- Géobios, Villeurbanne, vol. 30, p. 651-672.
A., P. & D. (2002).- Parallel biozonation in the Upper Callovian and the Lower Oxfordian based on the Peltoceratinae subfamily (Ammonitina, Aspidoceratidae).- Abhandlungen der Geologischen Bundesanstalt, Wien, vol. 57, p. 501-507.
S. (2008).- Cyclostratigraphie des séries sédimentaires du Jurassique supérieur (Sud-Est de la France, Nord de la Tunisie) : contrôle astro-climatique, implications géochronologiques et séquentielles.- Thèse de Doctorat, Université Pierre et Marie Curie, Paris, 313 p.
S., L.A., E., P.Y., B., D., D. & J. (2008).- Astronomical calibration of the Early Oxfordian (Vocontian and Paris basins, France): consequences of revising the Late Jurassic time scale.- Earth and Planetary Science Letters, Amsterdam, vol. 276, p. 1-2 & 40-51.
J.H. & J.C.W. (1993).- The Jurassic geology of Dorset: guide to excursions.- International symposium (September 1993), University College London, p. 42-48.
E., R., F., P., D. & M. (1997).- Oxfordien. In: E. & P. (coord.), Biostratigraphie du Jurassique ouest européen et méditerranéen : zonations parallèles et distribution des invertébrés et microfossiles.- Bulletin des Centres de Recherches Exploration-Production elf aquitaine, Pau, Mémoire 17, p. 79-86 & 144-147.
N.D. (1996).- The rediscovery of Creniceras renggeri () in the Upper Oxford Clay near Weymouth.- Proceedings of the Dorset Natural History and Archaeological Society, Dorchester, vol. 117, p. 153-154.
N.D. (1997).- Ammonites from the Oxford Clay near Budmouth School and Tidmoor Point, Weymouth, and their bearing on the Calllovian/Oxfordian boundary.- Proceedings of the Dorset Natural History and Archaeological Society, Dorchester, vol. 119, p. 117-127.
N.D. (1999).- Ammonite assemblages of the Upper Oxford Clay (Mariae Zone) near Weymouth.- Proceedings of the Dorset Natural History and Archaeological Society, Dorchester, vol. 121, p. 77-100.
G., D. & D. (1994).- Tectonique syn-sédimentaire et variations du niveau marin pendant le dépôt de la formation des Terres noires (Callovien supérieur - Oxfordien moyen ; Bassin du Sud-Est, France).- Comptes-Rendus de l'Académie des Sciences de Paris, (Série II), vol. 319, p. 559-565.
G. & D. (1979).- Les influences mésogéennes et boréales dans le Dogger et le Malm (Oxfordien) des Alpes-Maritimes.- 7e Réunion annuelle des Sciences de la Terre, Lyon, p. 144.
D. (1989).- Les "Terres noires" d'âge Callovien supérieur à Oxfordien moyen des chaînes sub-alpines du Sud (Diois, Baronnies, Dévoluy) : nouvelles données biostratigraphiques.- Comptes-Rendus de l'Académie des Sciences de Paris, (Série II), vol. 308, p. 531-536.
D. & D. (1994a).- Nouvelles unités biochronologiques de la zone à Mariae (Oxfordien inférieur).- Géobios, Villeurbanne, Mémoire spécial 17, p. 203-209.
D. & D. (1994b).- The Callovian-Oxfordian Boundary in the Basin of South of France. In: F. (ed.), 4th Oxfordian and Kimmeridgian working groups meeting in the South-Eastern France Basin.- Guidebook, Lyon, p. 24-26.
D. & D. (1994c).- The Savournon section: Upper Callovian (Lamberti zone) to Lower Oxfordian (Mariae zone) under "Terres Noires" facies. In: F. (ed.), 4th Oxfordian and Kimmeridgian working groups meeting in the South-Eastern France Basin.- Guidebook, Lyon, p. 95-99.
D. & D. (1994d).- The Thuoux section: Callovian-Oxfordian boundary (Lamberti to Mariae zone) under "Terres Noires" facies. In: F. (ed.), 4th Oxfordian and Kimmeridgian working groups meeting in the South-Eastern France Basin.- Guidebook, Lyon, p. 103-106.
D., D. & A. (1997).- Ammonites et limite Callovien-Oxfordien dans les "Terre Noires" du Diois (Bassin du Sud-Est, France) : exemples des coupes de Thuoux et Savournon.- Géobios, Villeurbanne, vol. 30, p. 519-540.
D.B., D.N. & M.A. (2002).- Biostratigraphy of the Upper Boreal Bathonian and Callovian of European Russia.- 6th International Symposium on the Jurassic System, Palermo, Abstract, p. 81-82.
R.A. (1990).- The Oxfordian ammonite succession near Liesberg BE and Péry BE, northern Switzerland.- Eclogae geologica Helvetia, Basel, vol. 83, p. 177-199.
A. & M. (1995).- Ammoniten aus dem Callovium/Oxfordium-Grenzbereich von Nordchile.- Berliner Geowissenschaft Abhandlungen A, vol. 169, p. 1-40.
R. (2010).- The evolution of ammonite associations during the Early Oxfordian (Mariae and Cordatum zones) in the Jura area (Eastern France).- Carnets de Géologie [Notebooks on Geology], Brest, Article 2010/07 (CG2010_A07), 15 p.
A. (1955).- Die Eisen- und Manganerze der Schweiz. Stratigraphie und Palaeontologie des oolithischen Eisenerzlagers von Herznach und seiner Umgebund.- Beiträge zur Geologie der Schweiz, Bern, (Geotechnische Serie), Lieferung XIII, Bd. 5, 238 p. (107 Taf.)
V.G. (1975).- Ammonites and zonal stratigraphy of the Lower Oxfordian of North Siberia.- Trudy Institut Geologiya y Geofisika Akademiya Nauk SSSR Sibirskikh Otdelemia, Novosibirsk, vol. 275, 139 p. [in Russian]
D.N. & M.A. (2004).- Preliminary report on the Callovian/ Oxfordian boundary in the Central Russia.- 32nd International Geological Congress, Abstract vol. 1 (177-19), p. 824.
D.N. & M. (2005).- Infrazonal stratigraphy and ammonites of the Middle-Upper Jurassic boundary beds of the European Russia (in Russian). In: V.A., M.A. & O.S. (eds.), Materials of the first All-Russian Meeting "Jurassic System of Russia: problems of stratigraphy and paleogeography".- Moscow: GIN RAS, p. 128-134.
D.N., M.A., A., M., E. & O. (2006).- Dubki (Saratov region, Russia): the reference section for the Callovian/Oxfordian boundary.- Proceedings of the 7th international Symposium on Jurassic Stratigraphy, Krakow (2006), Abstract vol., p. 36-38.
W. (1973).- Ammoniten und Ostreen (Biostratigraphie, Ökologie, Zoogeographie) des Callovium/Oxfordium-Grenzbereichs im Wiehengebirge.- Münstersche Forschungen zur Geologie und Paläontologie, vol. 27, 209 p. (25 pls.)
V. (1938).- Contribution à la connaissance des Cardioceratidés.- Mémoires de la Société géologique de France, Paris, (nouvelle série), n° 34, 134 p. (20 pls.)
D. (1979).- Un nouvel horizon paléontologique : l'horizon à Paucicostatum (Oxfordien inférieur, zone à Mariae, base de la sous-zone à Scarburgense).- Comptes-Rendus sommaires de la Société géologique de France, vol. 3, p. 122-124.
D. (1984).- Ammonites et paléoenvironnements : une nouvelle approche.- Géobios, Villeurbanne, Mémoire spécial 8, p. 101-107.
D. (1986).- L'évolution des Cardioceratinae d'Europe occidentale dans leur contexte paléobiogéographique (Callovien supérieur – Oxfordien moyen).- Thèse de Doctorat, Université de Bourgogne, Dijon, 395 p.
D. & G. (1979).- Colonisations faunistiques dans l'Oxfordien inférieur d'Europe occidentale. Exemple pris chez les Ammonoïdés.- 7e Réunion annuelle des Sciences de la Terre, Lyon, p. 311.
D., D., G., P.C. de & T. (1990).- Les peuplements d'ammonites du Bathonien supérieur à l'Oxfordien moyen dans les Baronnies (Bassin du Sud-Est, France) : comparaisons avec la plate-forme nord-européenne.- Bulletin des Centres de Recherches Exploration-Production elf aquitaine, Pau, vol. 14, n° 2, p. 465-479.
D., R. A., A. & D. (2000).- Les ammonites du Callovien terminal (zone à Lamberti) et de l'Oxfordien basal (zone à Mariae) d'Argovie (Suisse septentrionale).- Revue Paléobiologique, Genève, vol. 19, n° 1, p. 179-189.
G. (1996).- Oxfordian Working Group.- International Subcommission on Jurassic Stratigraphy, Newsletter, vol. 5, p. 29
G. (2003).- Oxfordian Working Group.- International Subcommission on Jurassic Stratigraphy, Newsletter, vol. 30, p. 16-18.
G. (2004).- Oxfordian Working Group.- International Subcommission on Jurassic Stratigraphy, Newsletter, vol. 31, p. 9-11.
G. (2006).- Oxfordian Working Group.- International Subcommission on Jurassic Stratigraphy, Newsletter, vol. 33, p. 16-19.
G., F. & K.N. (2007).- The cardioceratid succession and the recognition of the Callovian-Oxfordian boundary at Savournon (SE France).- XXIIIa Jornadas de la Sociedad Española de Paleontología, Caravaca de la Cruz, 2007, Libro de resumenes, p. 135-136.
N. & J.D. (1995).- Field guide to the Jurassic of the Isles of Raasay and Skye, Inner Hebrides, NW Scotland. In: P. D. (ed.), Field geology of the British Jurassic.- The Geological Society, London, p. 209-280.
K.N. (1994a).- A review of the suitability of key British Callovian-Oxfordian and Oxfordian-Kimmeridgian sites as GSSP for stage boundaries (Abstract). In: F. (ed.), 4th Oxfordian and Kimmeridgian working groups meeting in the South-Eastern France Basin.- Guidebook, Lyon, p. 15-16.
K.N. (1994b).- The Callovian-Oxfordian boundary in Britain: a review of key sections and their correlation with the proposal Global Stratotype Section and Point for the Oxfordian in Haute Provence, France.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 110, p. 201-208.
K.N., G., M.B., G.D., J.K., P. & J. (2006).- Integrated stratigraphical study of the candidate Oxfordian Global Stratotype Section and Point (GSSP) at Redcliff Point, Weymouth, Dorset, UK.- Volumina Jurassica, Warsaw, vol. 6, p. 1-13.
K.N., G. & J.K. (2009a).- The ammonite faunas of the Callovian-Oxfordian boundary interval in Europe and their relevance to the establishment of an Oxfordian GSSP.- Volumina Jurassica, Warsaw, vol. 7, p. 89-99.
K.N., G., M.B., G.D., J.K., P. & J. (2009b).- Integrated stratigraphical study of the candidate Oxfordian Global Stratotype Section and Point (GSSP) at Redcliff Point, Weymouth, Dorset, UK.- Volumina Jurassica, Warsaw, vol. 7, p. 101-111.
H. (1998).- Upper Callovian to upper Oxfordian Ammonite Biostratigraphy of the transect Chacay Melehue-Sierra de Reyes, Argentina.- Cuadernos de Geología Ibérica, Madrid, vol. 24, p. 261-275.
H. (2006).- Oxfordian and late Callovian ammonite faunas and biostratigraphy of the Neuquén-Mendoza and Tarapaca basins (Jurassic, Ammonoidea, Western South-America).- Boletin del Instituto de Fisiografía y Geología, Rosario, vol. 76, n° 1-2, p. 1-70.
N.E. & D. (1996).- Dinoflagellate cysts from two potential candidates for the Oxfordian stage basal boundary stratotype (GSSP), south-eastern France - a preliminary review.- Jurassic Microfossil Group, Newsletter, Copenhagen, vol. 5, p. 4-14.
P. (2009).- La Sous-Famille des Taramelliceratinae (Ammonitina, Haploceratoidea, Oppeliidae) de l'Oxfordien moyen et supérieur (Zone ŕ Plicatilis, Sous-Zone ŕ Vertebrale - Zone ŕ Bimammatum, Sous-Zone ŕ Berrense) du Nord de la Vienne, France (Province subméditerranéenne).- Carnets de Géologie [Notebooks on Geology], Brest, Brest, Mémoire 2009/02 (CG2009_M02), 101 p.
J., M.G., J.W., K.H., H.R., O. & (1996).- Revised guidelines for the establishment of global chronostratigraphic standards by the International Commission on Stratigraphy (ICS).- Episodes, Ottawa, vol. 19, p. 77-81.
J.B. Jr. (1919).- Some American Jurassic Ammonites of the Genera Quenstedticeras, Cardioceras and Amoeboceras, Family Cardioceratidae.- United State Geological Survey, Professional Papers, Washington, vol. 118, 64 p. (24 pls.)
M.A. & E.U. (2003).- Polymorphism in the some Early Oxfordian oppeliids (Ammonoidea) of the Russian Platform. In: N.A., T.I., V.E. et al. (eds.), Modern questions of geology.- Materials of the 3rd Lectures in Memory of , Moscow, vol. 26-28, p. 245-248.
R.M. (1975).- The stratigraphy of the Callovian and Oxfordian stages (Middle to Upper Jurassic) in northern Scotland.- Scottish Journal of Geology, Edinburgh, vol. 11, p. 51-78.
E.M. (2008).- Late Callovian and Early Oxfordian ostracods from the Dubki section (Saratov area, Russia): implications for stratigraphy, paleoecology, eustatic cycles and palaeobiogeography.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 249, p. 25-45.
J., E., S., C., D. & M. (1997).- Callovien. In: E. & P. (coord.), Biostratigraphie du Jurassique ouest européen et méditerranéen : zonations parallèles et distribution des invertébrés et microfossiles.- Bulletin des Centres de Recherches Exploration-Production elf aquitaine, Pau, Mémoire 17, p. 63-78 & 140-143.
J., D., D., A. & R. (2006).- Les ammonites du Callovo-Oxfordien des sondages ANDRA dans l'Est du Bassin de Paris : synthèse bio-chronostratigraphique, intérêts paléoécologique et paléobio-géographique.- Comptes-Rendus des Geosciences, Paris, vol. 338, p. 834-853.
J. (1966).- The Oxford Clay of Skye, Scalpay and Eigg.- Scottish Journal of Geology, Edinburgh, vol. 2, p. 243-252.
J.P., D., A. & D. (1993).- Le Callovien et l'Oxfordien p.p. du Boulonnais : nouvelles données biostratigraphiques.- Acta Geologica Polonica, Warszawa, vol. 43, n° 3-4, p. 169-182.